Exploring the SEC Analysis of CRISPR Molecules and Their Complexes
Abstract
Size Exclusion Chromatography (SEC) is a powerful separation technique for protein analysis, though it can show its limitations when there are protein-to-hardware or protein-to-packing material interactions. CRISPR Cas9 proteins were confirmed in this work to be a challenging sample with which to work. Significant adsorptive losses were observed when using conventional stainless-steel hardware columns, suggesting that extensive method development would be required to achieve satisfactory separations. Recent advances in SEC column technology, including hydrophilic MaxPeak™ High Performance Surfaces (h-HPS) modified hardware surfaces, has provided significant reductions in undesired secondary interactions such that there has become possible to separate proteins with PBS based buffers at or near physiological pH (about 7.4) and with relatively low ionic strengths (about 150 to 300 mM). Leveraging this technology, we have demonstrated the use of MaxPeak Premier Protein SEC 250 Å columns to obtain improved SEC-UV recoveries of Cas9 and to begin investigating ribonucleoprotein (RNP) complex formation at varied stoichiometric ratios. Separately, volatile ammonium acetate mobile phases to make a direct native MS-based observation of a ~160 kDa mass that corresponds the intact mass of the free Streptococcus Cas9 protein. With these preliminary method insights, it is believed that even more success might be found in the SEC analysis of other CRISPR molecules, such as smaller ~124 kDa Staphylococcus aureus derived Cas9 homologs.
Benefits
- MaxPeak Premier Protein SEC 250 Å Column offers exemplary Cas9 recoveries due to improved column surface inertness
- Method considerations for SEC-UV of Cas9, gRNA, and RNP Complexes
- Preliminary insights on SEC-MS of Cas9
Introduction
Clustered regularly interspaced short palindromic repeats (CRISPR) was coined by Francisco Mojica in 1993.1 CRISPR is a gene editing technique that requires two components: a CRISPR associated (Cas) protein enzyme and a guide RNA molecule. In recent years, CRISPR has been successfully applied to gene editing in foods, plants, biofuel production, and now in vitro gene edit of human cells and in vivo gene editing in human patients.1 Even more recently, Professors Jennifer Doudna and Emmanuelle Charpentier were awarded a Nobel Prize for their contributions to the discovery and use of the CRISPR system. In some of their most noteworthy work, they used E. coli expressed, recombinant Cas9 (from Streptococcus pyogenes) together with crispr RNA (crRNA) and an in vitro transcribed trans-activating crRNA (tracrRNA or trRNA). With this, they demonstrated that a complex formed from these molecules could cut purified DNA and thereby facilitate gene editing (Jinek et al., 2012).2 Moreover, they showed that target site specificity could be programmed with custom-designed crRNAs and that the trRNA and crRNA could be fused into a single-guide RNA (sgRNA). The concept of sgRNAs has now become widely applied in genome editing.1
Altering genomes in living cells is not a novel concept and had been revolutionized in the late 1980s but shown to be relatively inefficient. Genome-editing technologies such as designer zinc fingers (ZFs)1–4, transcription activator–like effectors (TALEs)4–6 and homing meganucleases5 were each on their own major breakthroughs in gene editing technologies. The CRISPR technology has shown to be more precise in executing double-stranded DNA breaks at precise DNA loci and are scalable, affordable, and easy to engineer.3-4 Therapeutic applications are now emerging. Indeed, the U.S. Food and Drug Administration (FDA) granted Rare Pediatric Disease designation to EDIT-301 by Editas Medicine for an investigational, CRISPR gene-edited medicine for the treatment of sickle cell disease (SCD) and transfusion-dependent beta thalassemia (TDT). Also, currently in a Phase III clinical trial is CTX001 by Vertex Pharmaceutical which is a patient-derived CRISPR-Cas9-edited stem cell therapy for Beta Thalassemia.5-6
CRISPR associated protein 9 (Cas9) is responsible for locating and cleaving target DNA, both in natural and in artificial CRISPR/Cas systems. Type II Cas9 protein reported here is a dual RNA-guided DNA endonuclease enzyme associated with the CRISPR adaptive immune system in Streptococcus pyogenes.2,6 When this ~160 kDa Cas9 protein combines with crRNA and trRNA or single guide-RNA to form the RNP complex for gene-editing, it permits specific cutting of the DNA. The intermolecular interactions and design of an RNP for therapeutic gene edits are very important. Without careful design and study, unwanted off-target cleavages could occur at unspecified loci in a cell’s genome. SEC technique has been proven to be an effective technique to study the biophysical properties of proteins and their complexes. One of the major limitations in protein SEC is the presence of undesired electrostatic and hydrophobic secondary interactions. Some proteins are particularly prone to these complications. Cas9 proteins have been thought to be difficult to analyze by SEC. An analysis of its crystal structure presents a hypothesis as to why this might be (Figure 1). With the crystal structure for S. pyogenes Cas9, Shenal, and co-workers observed that the protein has a highly basic cleft to facilitate the binding of its complementary nucleic acids. Lys and Arg resides are highlighted in blue to help illustrate the location of this cleft.7 Despite this, Streptococcus Cas9 also presents and acidic domain. The electronegativity of this motif might explain why some have reported challenges in being able to chromatograph this protein. The strong electronegativity corresponding to a concentrated region of Asp and Glu residues would be predicted to show affinity for and adsorptive losses from metallic column hardware.
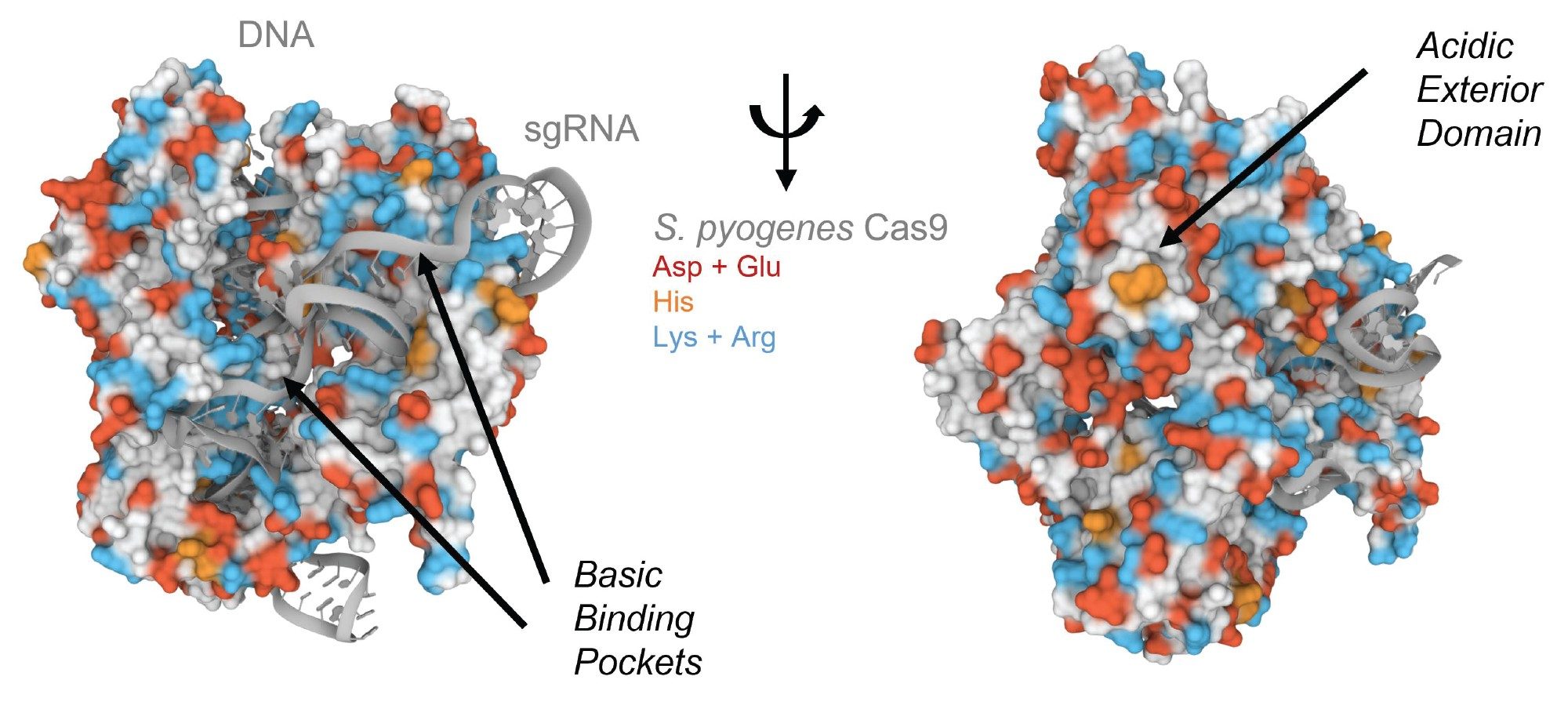 Figure 1. Crystal structure of S. pyogenes Cas9 in complex with a guideRNA and target DNA molecule. Protein residues are displayed as a surface representation and the nucleic acids are shown as cartoons. Aspartic acid and glutamic acid residues are highlighted in red and histidine in orange while lysine and arginine are highlighted in blue. Images were created using Mol* and RCSB PDB ID 4OO8. (D. Sehnal, S. Bittrich, M. Deshpande, R. Svobodová, K. Berka, V. Bazgier, S. Velankar, S.K. Burley, J. Koča, A.S. Rose (2021) Mol* Viewer: modern web app for 3D visualization and analysis of large biomolecular structures. Nucleic Acids Research. doi:10.1093/nar/gkab31.7
Figure 1. Crystal structure of S. pyogenes Cas9 in complex with a guideRNA and target DNA molecule. Protein residues are displayed as a surface representation and the nucleic acids are shown as cartoons. Aspartic acid and glutamic acid residues are highlighted in red and histidine in orange while lysine and arginine are highlighted in blue. Images were created using Mol* and RCSB PDB ID 4OO8. (D. Sehnal, S. Bittrich, M. Deshpande, R. Svobodová, K. Berka, V. Bazgier, S. Velankar, S.K. Burley, J. Koča, A.S. Rose (2021) Mol* Viewer: modern web app for 3D visualization and analysis of large biomolecular structures. Nucleic Acids Research. doi:10.1093/nar/gkab31.7
In this application note, we demonstrate simple SEC methods that could be considered for the characterization of Cas proteins, CRISPR RNA and the RNP complexes that they form. Here, a newly developed SEC packing material based on 250 Å BEH™ particles and a hydroxy PEO bonding was applied along with hardware modified with hydrophilic MaxPeak High Performance Surfaces (h-HPS). The h-HPS surface hardware showed exemplary recoveries of multiple Cas9 molecules when compared to a conventional stainless-steel column hardware. That recoveries of Cas9 nucleases are so significantly improved with h-HPS hardware bodes well for more opportunities to come from continued method development and the application of SEC CRISPR molecule characterization, from drug design to formulation and stability studies. In one case, a phosphate buffered saline based mobile phase has been successfully applied and used to study individual injections as well as ratio metric mixtures of Cas9 with RNA. Indications of an equilibrium-based formation of RNP complexes has been observed. Separately, volatile ammonium acetate mobile phases have been applied along with QTof mass spectrometry to make a direct native mass spectrometry (MS)-based observation of a ~160 kDa mass that corresponds the intact mass of a free Streptococcus Cas9 protein. With these preliminary method insights, it is believed that even more success might soon be found in the SEC analysis of CRISPR molecules, whether they be SEC used with multidimensional separations, SEC applied for native MS, or the use of these tools for additional investigations into the smaller, and perhaps more tractable, Staphylococcus aureus derived Cas9 homologs (~124 kDa).
Experimental
Sample Description
Cas9 Nuclease Protein NLS acquired from Dharmacon/Horizon and contains the Streptococcus pyogenes Cas9 (Csn1) gene, p/n: CAS12206 in a concentration of 10 µg/µL and diluted with milli-Q water to 5 µg/µL. Milli-Q water from Millipore. 20 mM Phosphate, 276 mM NaCl, 5.4 mM KCl pH 7.4 (2x PBS buffer) prepared by adding 2 packets PBS obtained from Sigma, p/n: P3813 to 1 liter of Milli-Q water and filtered prior to use through a sterile 0.2 µm nylon filter (Thermo Scientific p/n: 1630020). 5-fold dilution of IonHance™ CX-MS Concentrate B to make 320 mM Ammonium Acetate, pH 8.5 in 4% ACN.
RNP Complex Formation
RNP complex was formed by initially incubating sgRNA, Edit-R Synthetic sgRNA Non-targeting Control #1, p/n: U-009501-01-05 acquired from Dharmacon/Horizon at 60 °C for ten minutes and allowing to cool to room temperature for fifteen minutes. A 1:1 and a 3:1 RNP complex mixtures of heated sgRNA to Cas9 Nuclease Protein NLS were then prepared and mixed gently by vortex. Mixture was then centrifuged at 8000 rpm for thirty seconds and transfer to a Waters polypropylene 12 x 32 mm screw neck vial, 300 µL volume (p/n: 186002639).
LC Conditions
|
LC system: |
ACQUITY™ UPLC™ H-Class Bio |
|
Detection: |
ACQUITY UPLC TUV Detector (Titanium Flow Cell, 5 mm, 1500 nL) |
|
Wavelength: |
280 nm (Cas9 samples) and 260 nm (RNP Formation) |
|
Data acquisition: |
Empower™ Pro 3 Feature Release 3 |
|
Vials: |
Max Recovery Vials and Caps (Waters p/n: 186000327C) and Waters 300 μL polypropylene screw neck vial (Waters p/n: 186004112) |
|
Column(s): |
XBridge™ Premier Protein SEC 250 Å, 2.5 µm, 4.6 x 150 mm ACQUITY Premier Protein SEC 250 Å, 1.7 µm, 4.6 x 300 mm |
|
Column temperature: |
35 °C |
|
Sample temperature: |
8 °C |
|
Sample manager washes: |
18.2 MΩ water |
|
Seal wash: |
10% HPLC grade Methanol/90% 18.2 MΩ water (v/v) |
|
Injection volume: |
gRNA (2 µL of 5 µg/µL); RNP Formation (2 µL of 1:1 and 3:1 complex) |
|
Flow rate: |
0.2 mL/min |
|
Mobile phase A: |
2X PBS: Phosphate Buffered Saline (20 mM Phosphate, 276 mM NaCl, 5.4 mM KCl pH 7.4) |
|
Sample(s): |
Cas9 Nuclease Protein NLS |
|
Gradient: |
Isocratic |
SEC-MS
|
LC system: |
ACQUITY Premier Binary Solvent Manager |
|
Detection: |
ACQUITY UPLC TUV Detector (Titanium Flow Cell, 5 mm, 1500 nL) |
|
Wavelength: |
280 nm |
|
Data acquisition |
MassLynx™ v4.1 |
|
Vials: |
Max Recovery Vials and Caps (Waters p/n: 186000327C) and Waters 300 μL polypropylene screw neck vial (Waters p/n: 186004112) |
|
Column(s): |
ACQUITY Premier Protein SEC 250 Å, 1.7 µm, 4.6 x 150 mm |
|
Column temperature: |
30 °C |
|
Sample temperature: |
8 °C |
|
Seal wash and FTN SM washes: |
10% HPLC grade Methanol/90% 18.2 MΩ water (v/v) |
|
Injection volume: |
SpCas9 (10 µL) |
|
Flow rate: |
0.1 mL/min |
|
Mobile phase A: |
5-fold Diluted IonHance CX-MS Concentrate B to make 320 mM Ammonium Acetate, pH 8.5 in 4% ACN |
|
Sample(s): |
Cas9 Nuclease Protein NLS |
|
MS system: |
Xevo™ G2-XS QToF |
|
Quadrupole settings: |
1000/2000/3940 25%/25%/25%/25% |
|
Acquisition window: |
2000 to 7000 m/z |
|
Capillary: |
3 kV |
|
Scan rate: |
2 Hz |
|
Source temperature: |
100 °C |
|
Desolvation gas flow: |
600 L/Hr |
|
Desolvation gas temperature: |
450 °C |
|
Cone gas flow: |
50 L/Hr |
Results and Discussion
Previous work has explored the benefits of a novel 250 Å BEH HO-PEO bonded particle and MaxPeak h-HPS Hardware for mitigating electrostatic and hydrophobic secondary interactions during SEC.8 In this work, we showed it to be possible to apply a platform method and mobile based on a 2x concentration strength of phosphate buffered saline (20 mM Phosphate, 276 mM NaCl, 5.4 mM KCl pH 7.4; 2x PBS). This served as a starting point for investigations into the chromatography of a Streptococcus pyogenes Cas9. Experiments were first performed to compare the peak shape and recovery of SEC columns prepared with Protein SEC 250 Å packing material and standard metallic hardware versus h-HPS modified hardware. Specifically, we compared the performance of conventional stainless-steel column hardware to that of h-HPS column hardware using 4.6 x 300 mm dimensions and the same packing hydroxy-terminated BEH 250 Å, 2.5 µm packing material. Figure 2 displays the chromatograms obtained. As predicted, the highly concentrated electrostatic binding sites of a typical CRISPR Cas9 protein appears to have led to adsorptive losses on the conventional stainless-steel hardware.
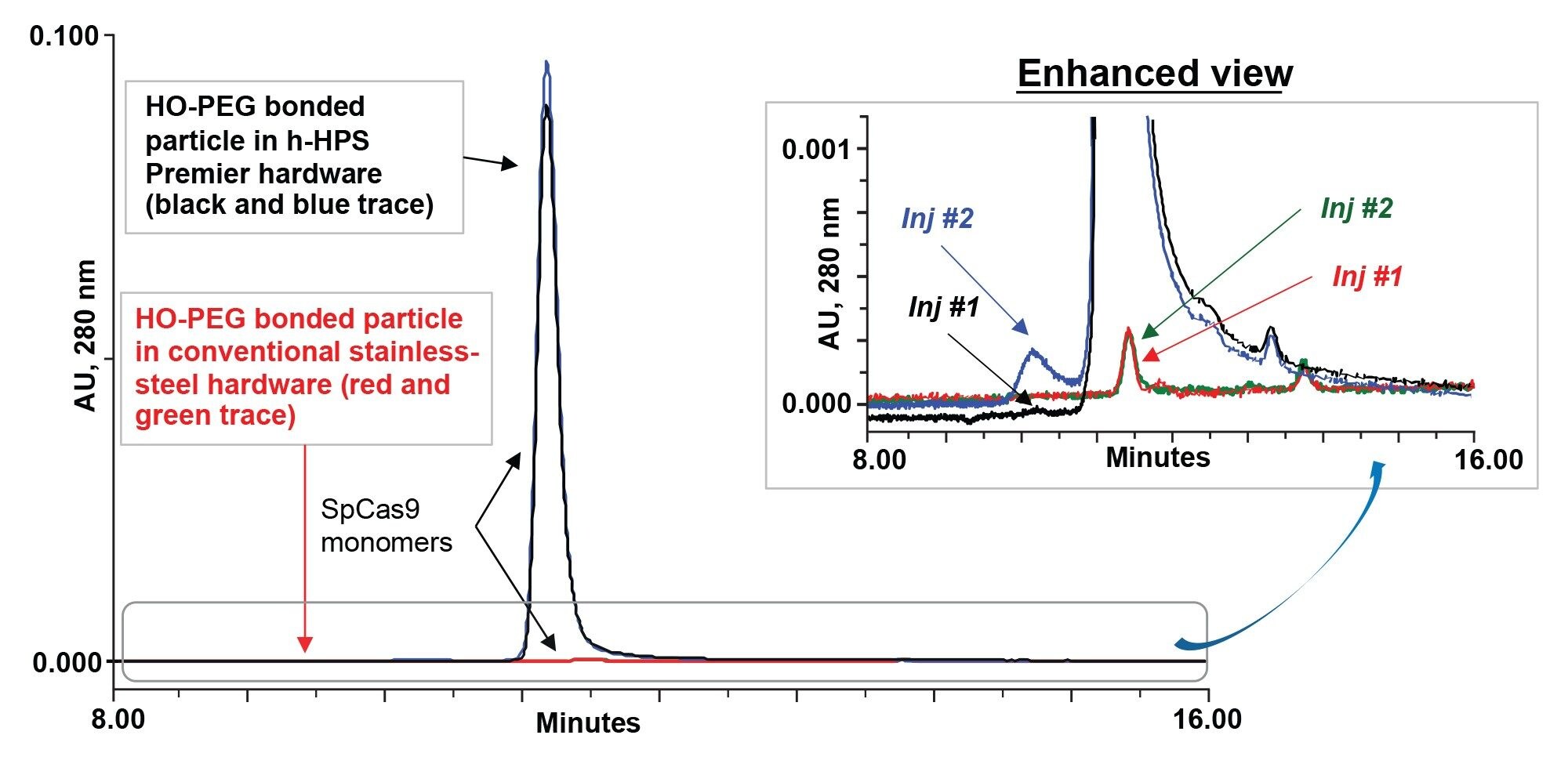 Figures 2. Comparison of SpCas9 proteins with conventional stainless-steel column hardware and hydrophilic MaxPeak High Performance Surfaces column hardware with 250 Å BEH particle with high coverage hydroxy-terminated PEO bonding using 2x PBS mobile phase. SpCas9 nuclease protein from S. pyogenes Cas9 exhibits strong electrostatic interactions with conventional stainless-steel hardware (red and green traces) for injections 1 and 2, respectively than h-HPS column hardware (black and blue traces) for injections 1 and 2, respectively.
Figures 2. Comparison of SpCas9 proteins with conventional stainless-steel column hardware and hydrophilic MaxPeak High Performance Surfaces column hardware with 250 Å BEH particle with high coverage hydroxy-terminated PEO bonding using 2x PBS mobile phase. SpCas9 nuclease protein from S. pyogenes Cas9 exhibits strong electrostatic interactions with conventional stainless-steel hardware (red and green traces) for injections 1 and 2, respectively than h-HPS column hardware (black and blue traces) for injections 1 and 2, respectively.
There was a sizable difference in the performance between conventional stainless-steel and h-HPS hardware. In Figure 2, h-HPS column hardware with BEH-PEO particle significantly minimized electrostatic properties of SpCas9 (Figure 1). Use of conventional stainless-steel column hardware made it near impossible to recover SpCas9. It is reasonable to suggest that the electrostatic domains of SpCas9 strongly adsorbed to exposed metallic hardware surfaces within the column. Even with h-HPS hardware, there was a small indication of conditioning; that is, aggregate recovery increased upon the second versus first injection (blue trace). Nevertheless, the first injection on the h-HPS column produced excellent recovery of the monomeric SpCas9 species. The stability effect of SpCas9 has not been examined to fully understand how quickly aggregates are formed.
Given its h-HPS hardware, the XBridge™ Premier Protein SEC 250 Å, 2.5 µm Column showed promising performance such that this type of column technology was selected for additional work on Cas9 proteins and an investigation into the formation and chromatographic behavior of RNP complexes.
RNP Complex Formation
The conformational changes in Cas9 are not completely understood, but Jinek and colleagues hypothesize that steric interactions or weak binding between protein side chains and RNA bases may induce changes.10 During formation of the RNP complex, it is essential to quickly assess the stability of compounds during and after complexation. To begin doing this by SEC, we used a ACQUITY Premier Protein SEC 250 Å 1.7 µm, 4.6 x 300 mm Column, and we studied the formation of RNPs using easy to prepare phosphate buffered saline as mobile phase. Figure 3a–d below shows SEC-UV traces for individual sgRNA, individual SpCas9, and RNP complexes resulting from 1:1 and 3:1 mixtures of sgRNA and SpCas9, respectively.
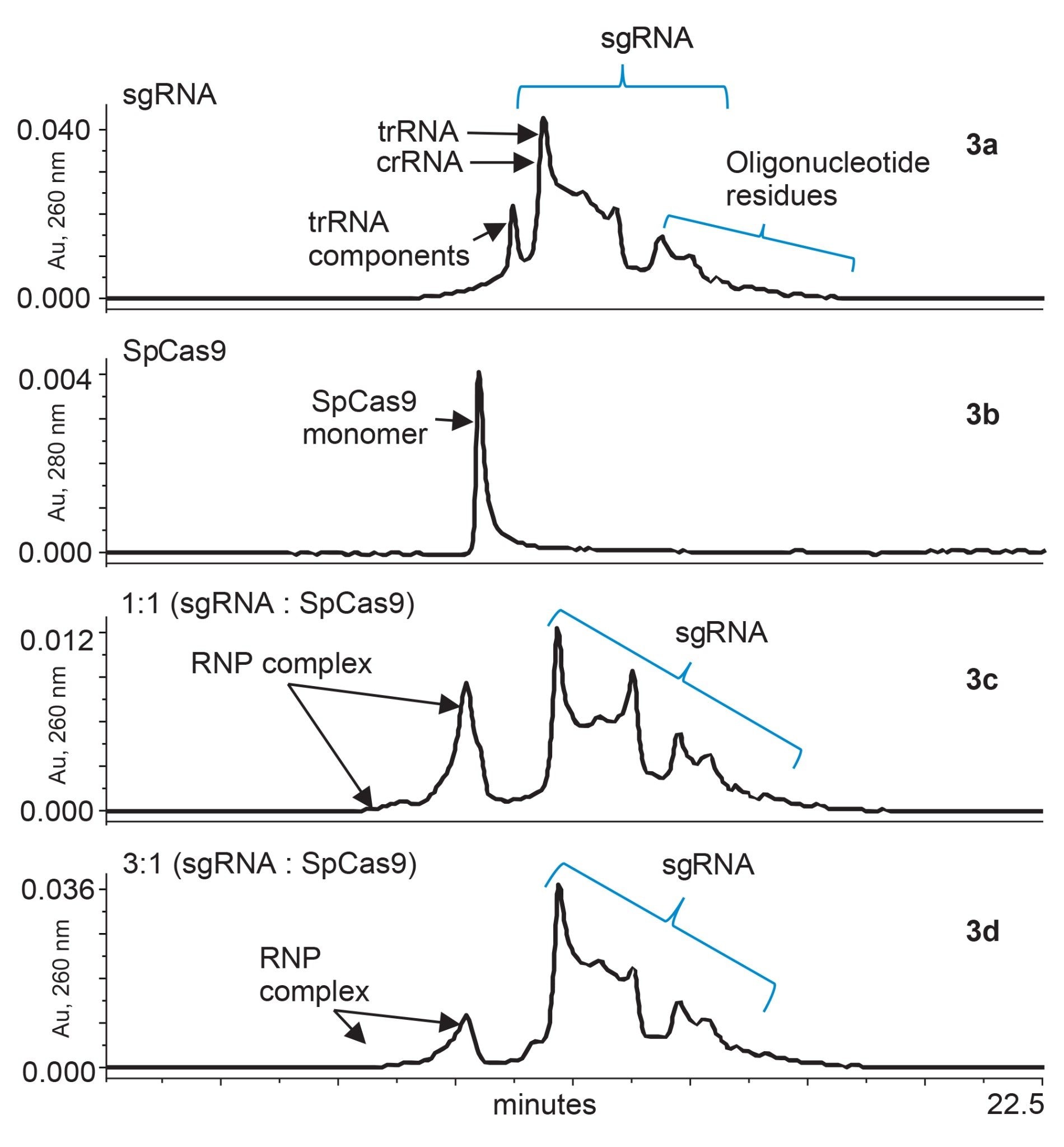 Figure 3a–d. Formation of RNP complex and individual guide-RNA and Cas9 protein were acquired using ACQUITY Premier Protein SEC 250 Å 1.7 µm 4.6 x 300 mm Column. Figure 3a UV chromatogram of sgRNA acquired at 260 nm. Fig 3b is Cas9 from s. pyogenes acquired at 280 nm. Figure 3c shows formation of RNP complex when sgRNA and Cas9 were combine at a 1:1 ratio. Figure 3d shows formation of RNP complex when sgRNA and Cas9 were combined at a 3:1 ratio.
Figure 3a–d. Formation of RNP complex and individual guide-RNA and Cas9 protein were acquired using ACQUITY Premier Protein SEC 250 Å 1.7 µm 4.6 x 300 mm Column. Figure 3a UV chromatogram of sgRNA acquired at 260 nm. Fig 3b is Cas9 from s. pyogenes acquired at 280 nm. Figure 3c shows formation of RNP complex when sgRNA and Cas9 were combine at a 1:1 ratio. Figure 3d shows formation of RNP complex when sgRNA and Cas9 were combined at a 3:1 ratio.
Figure 3a presents an SEC-UV chromatogram of engineered sgRNA that has a molecular weight of ~32 kDa. The provided chromatogram was acquired with UV 260 nm detection to monitor nucleic acid elution more selectively. Meanwhile, Figure 3b presents an SEC-UV 280 nm chromatogram for SpCas9 and a corresponding monomeric peak eluting at 11.34 minutes. In Figure 3c, the result of a 1:1 mixture of sgRNA to SpCas9 is provided. RNP complexes were observed to be partially formed as evidenced by the presence of new peaks elution between 9.00 and 11.09 minutes. The shifted elution of these RNP peaks is presumably due to the increased hydrodynamic radii of the combined protein-RNA complexes. A small portion of SpCas9 protein was observed to be unbound at this ratio of 1:1 as observed by a shoulder on RNP complex peak. In Figure 3d, the ratio of the sgRNA was tripled to a 3:1 ratio. This increase appears to have produce an abundance of signal for free sgRNA. With the excess amount of sgRNA, there also appeared to be commensurate decrease in the unbound SpCas9 shoulder peak. Although there is still much to be understood, SEC-UV analysis can be implemented as a quick check on RNP complexation and as a tool to monitor its stability. By studying various ratios of sgRNA to Cas9 protein for RNP formation, an assessment of binding affinity can also be made, which can then be implemented for drug design experiments.
Preliminary Investigation into Volatile Mobile Phases and SEC-MS
For heightened characterization of a Cas protein and its complexes, it would be advantageous to simultaneously access mass spectrometric information for eluting analytes. Separations of SpCas9 were accordingly studied not only with the above phosphate buffered saline mobile phases but also ammonium acetate solutions that can be directly hyphenated with online MS detection. It is critically important that only trace metal, MS-certified mobile phase components be used in this type of work, since sodium and potassium adducts can quite easily make a negative impact on MS quality. IonHance ammonium acetate concentrates were applied for this reason and an experiment to study a series of different dilution factors was performed. UV chromatograms corresponding to the use of 100 mM, 200 mM, 240 mM, 280 mM, 300 mM, and 400 mM ammonium acetate are displayed in Figure 4. Given the complications encountered with SpCas9 even with the use of phosphate-based buffers, it was not surprising to find that a minimum of approximately 300 mM ammonium acetate was required to obtain a symmetrical monomer peak and high recovery. While not studied here, there is reason to believe that other pH titrated ammonium acetate solutions might be equally or better suited to this application.
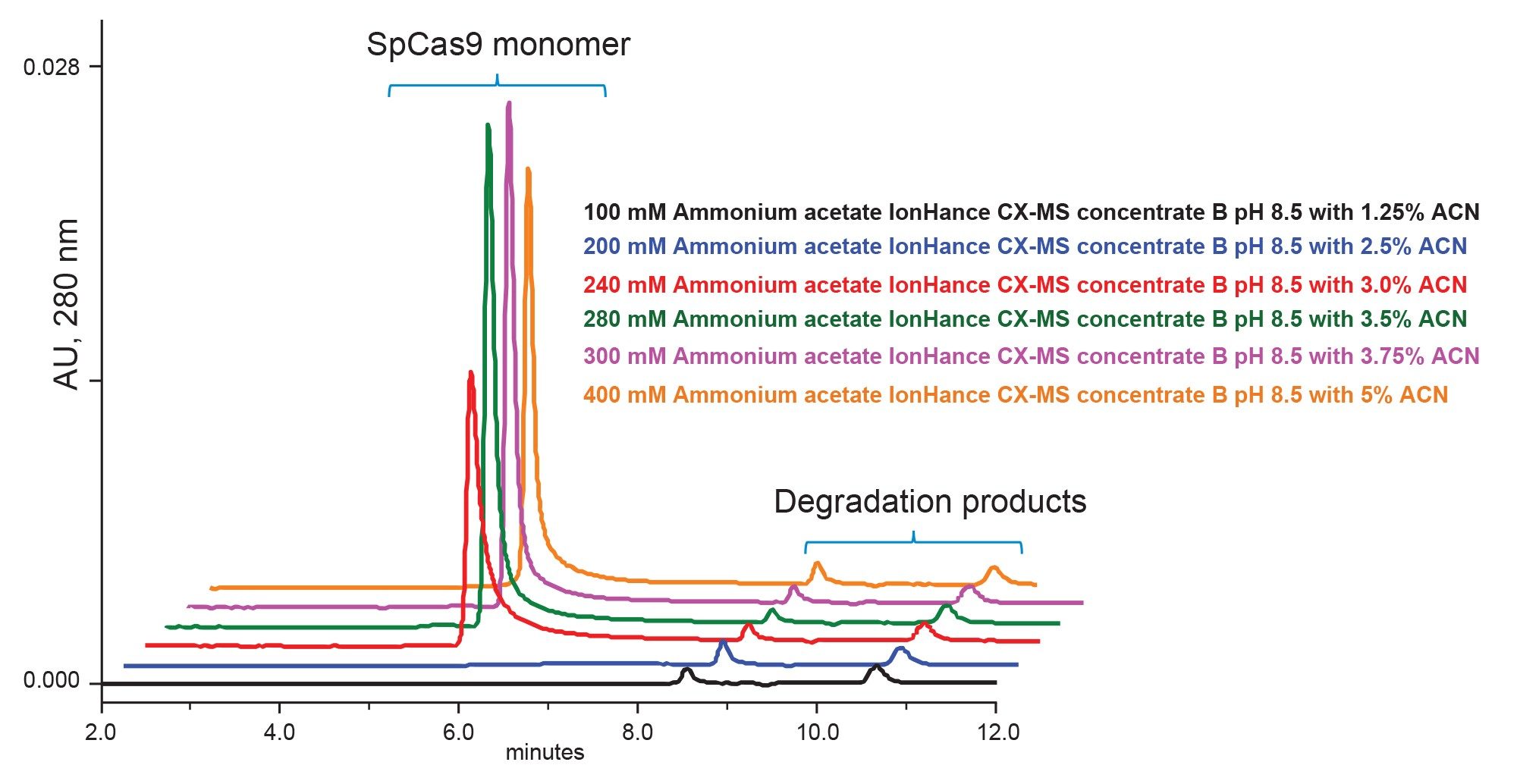 Figure 4. Analysis of SpCas9 using IonHance Concentrate B buffer diluted to make ammonium acetate pH 8.5 mobile phases comprised of 100 mM, 200 mM, 240 mM, 280 mM, 300 mM, and 400 mM.
Figure 4. Analysis of SpCas9 using IonHance Concentrate B buffer diluted to make ammonium acetate pH 8.5 mobile phases comprised of 100 mM, 200 mM, 240 mM, 280 mM, 300 mM, and 400 mM.
A 320 mM ammonium acetate mobile phase is relatively high for what is most frequently applied for native LC-MS. Nevertheless, we explored the optimization of QTof mass spectrometry for the detection of species in the high m/z range. Quadrupole mass filtering was adjusted to destabilize low m/z ions in exchange for more selective detection of 2000 m/z and above ions. A flow rate of 0.1 mL/min was employed to reduce the desolvation burden of the ESI source and, in turn, the UV and total ion chromatograms shown in Figure 5A were collected. The 320 mM ammonium acetate mobile phase used in this experiment produced excellent peak shapes for both the monomeric SpCas9 and its HMW species. A zoomed viewed of the chromatogram (shown in the inset) underscores the quality of this separation. SpCas9 is a large protein that is predicted to have a molecular weight of 158 kDa (UniProt Q99ZW2). It is also predicted to have multiple strong binding pockets for magnesium ions, and it is also plausible for it to be subject to post translational modifications. Moreover, the SpCas9 construct studied here was recombinantly prepared from the Csn1 gene with a combined C-terminal nuclear localization signal (NLS). This adds a stretch of basic residues with a combined mass of between 1 and 3 kDa. A summed raw mass spectrum was prepared from the monomer peak, and this is shown along with a MaxEnt1 deconvoluted spectrum in Figure 5B. Heterogeneity was observed in the raw spectrum which was appropriately carried through to the determination of the molecular weight information. The two most intense masses deconvoluted from this experiment was 161.5 and 161.9 kDa. Both of which are in reasonable agreement to the predicted average mass of the Csn1 gene product and a NLS C-terminal extension (weighing between 1 and 3 kDa). These results demonstrate that the MS-based interrogation of Cas9 proteins is at hand and that additional method development considerations for improved sensitivity and MS quality might provide the analytical capabilities needed to detect the complexation of a Cas9 protein to its guide RNA molecules. It should also be noted that even more success might be found in the SEC-MS analysis of smaller ~124 kDa Staphylococcus aureus Cas9 homologs.
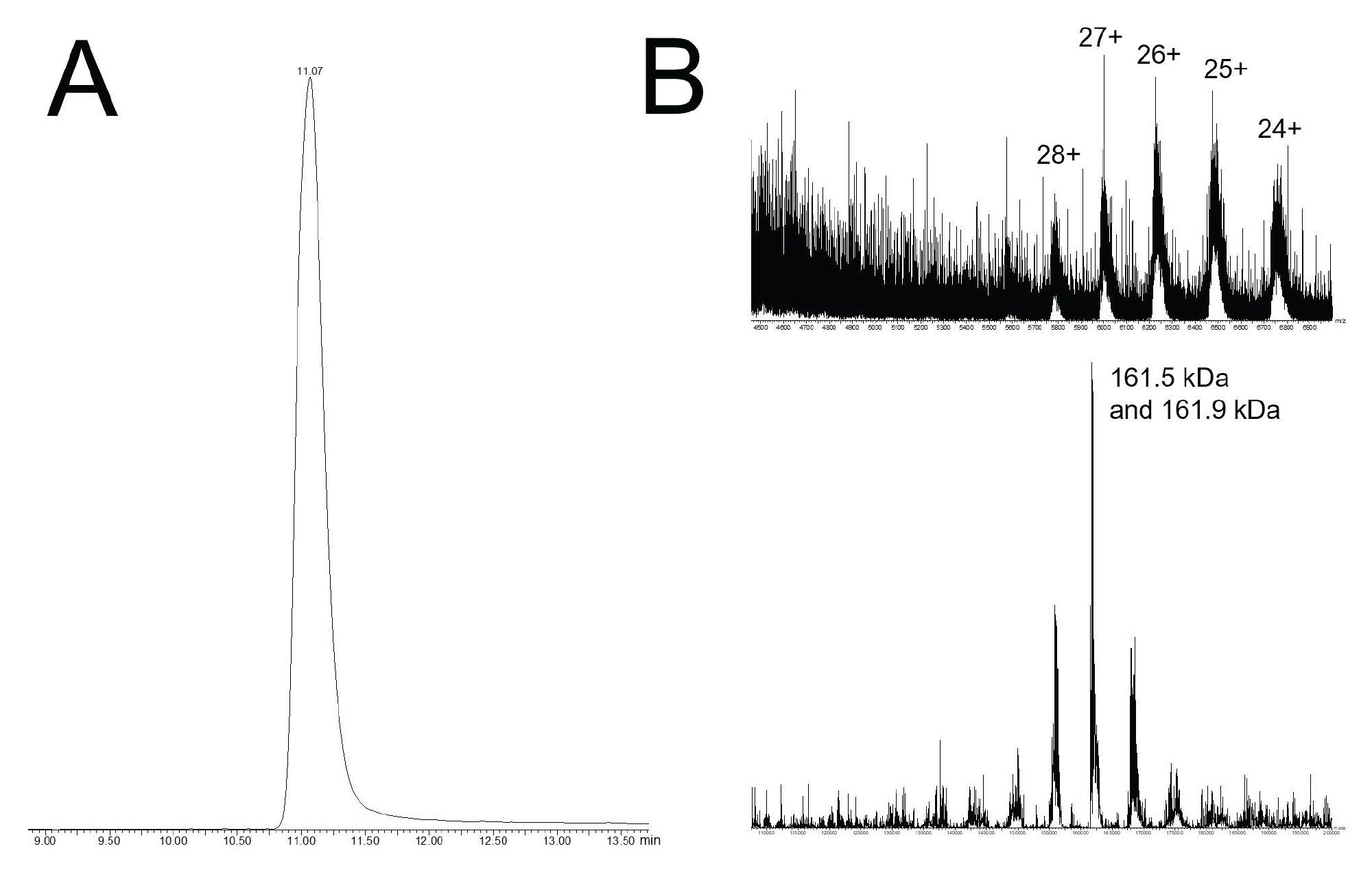 Figure 5. Native SEC-MS with a ACQUITY Premier Protein SEC 250 Å 1.7 µm 4.6 x 150 mm Column and a 320 mM pH 8.5 ammonium acetate mobile phase. (A) Total ion chromatogram acquired by QTof ESI-MS. (B) Summed raw spectrum corresponding to the monomer SpCas9 species and its corresponding MaxEnt1 deconvolution.
Figure 5. Native SEC-MS with a ACQUITY Premier Protein SEC 250 Å 1.7 µm 4.6 x 150 mm Column and a 320 mM pH 8.5 ammonium acetate mobile phase. (A) Total ion chromatogram acquired by QTof ESI-MS. (B) Summed raw spectrum corresponding to the monomer SpCas9 species and its corresponding MaxEnt1 deconvolution.
Conclusion
In this application note, we were able to create a simple SEC-UV method for analysis of a CRISPR/Cas9 protein and its RNP complexes. For this work, we employed Waters ACQUITY Premier Protein SEC 250 Å, 1.7 µm and XBridge Premier Protein SEC 250 Å, 2.5 µm Column technology which allowed the use of a simple PBS based mobile phase. It was previously demonstrated that the surface technologies in these new columns have significantly reduced nonspecific protein-column interactions.10 Electrostatic secondary interactions exhibited by Streptococcus pyogenes Cas9 proteins on conventional hardware columns were found to be greatly minimized in this work using the new h-HPS based column technology. This facilitated for more accurate SpCas9 analyses and afforded a chance to develop an online SEC-MS method using MS-certified mobile phase buffers comprised of IonHance ammonium acetate. Both SEC methods showed promise for the study and of CRISPR/Cas9, its counterpart nucleic acids, and the formation of RNP complexes.
References
- Lander Eric S (2016) The Heroes of CRISPR. Cell 164(1):18–28. doi.org/10.1016/j.cell.2015.12.041.
- Jinek M, Chylinski K, Fonfara I, Hauer M, Doudna JA, Charpentier E. A Programmable Dual-RNA-Guided DNA Endonuclease in Adaptive Bacterial Immunity. Science. 2012;337(6096):816–821. doi:10.1126/science.1225829.
- Le Cong, F. Ann Ran, David Cox, Shuailiang Lin, Robert Barretto, Naomi Habib, Patrick D. Hsu, Xuebing Wu, Wenyan Jiang, Luciano A. Marraffini, and Feng Zhang. Multiplex Genome Engineering Using CRISPR/Cas Systems. Science. 2013 February 15; 339(6121): 819–823. doi:10.1126/science.1231143.
- Prashant Mali, Luhan Yang, Kevin M. Esvelt, John Aach, Marc Guell, James E. DiCarlo, Julie E. Norville, and George M. Church. RNA-Guided Human Genome Engineering via Cas9. Science. 2013 February 15; 339(6121): 823–826. doi:10.1126/science.1232033.
- CRISPR Medicine News. News: Gene-Editing Clinical Trials for Beta Thalassemia - CRISPR Medicine (crisprmedicinenews.com).
- Deltcheva E, Chylinski K, Sharma CM, Gonzales K, Chao Y, Pirzada ZA, Eckert MR, Vogel J, Charpentier E. CRISPR RNA Maturation by trans-Encoded Small RNA and Host Factor RNase III. Nature. March 2011; 471(7340): 602–607. doi:10.1038/nature09886.
- David Sehnal, Sebastian Bittrich, Mandar Deshpande, Radka Svobodová, Karel Berka, Václav Bazgier, Sameer Velankar, Stephen K Burley, Jaroslav Koča, Alexander S Rose: Mol* Viewer: Modern Web App for 3D Visualization and Analysis of Large Biomolecular Structures, Nucleic Acids Research, 2021; 49(W1):431-437. doi:10.1093/nar/gkab31.
- Lavelay Kizekai, Stephen J. Shiner, Matthew A. Lauber. Waters ACQUITY and XBridge Premier Protein SEC 250Å Columns: A New Benchmark in Inert SEC Column Design. Waters Application Note, 720007493, 2022.
- Jinek M., Jiang F., Taylor D. W., Sternberg S. H., Kaya E., Ma E., Anders C., Hauer M., Zhou K., Lin S., Kaplan M., Iavarone A. T., Charpentier E., Nogales E., Doudna J. A., Structures of Cas9 Endonucleases Reveal RNA-Mediated Conformational Activation. Science. 2014; 343(6176). doi/10.1126/science.1247997.
- Stephan M. Koza, Hua Yang, and Ying Qing Yu. Modern Size-Exclusion Chromatography Separations of Biosimilar Antibodies at Physiological pH and Ionic Strength. Waters Application Note, 720007484, 2022.
720007727, September 2022