探索CRISPR分子及其复合物的SEC分析
摘要
体积排阻色谱(SEC)是一种用于蛋白质分析的强大的分离技术,尽管因蛋白质与硬件或蛋白质与填料的相互作用使其具有局限性。本研究证实,CRISPR Cas9蛋白质是一种棘手的样品。当使用传统的不锈钢硬件色谱柱时,观察到明显的吸附损失,这表明需要进一步开发方法才能达到令人满意的分离效果。SEC色谱柱技术的最新进展,包括亲水性MaxPeak™高性能表面(h-HPS)修饰的硬件表面,显著减少了不必要的次级相互作用,从而可以在生理pH值(约7.4)左右和相对较低的离子强度(约150~300 mM)下用PBS基缓冲液分离蛋白质。利用这项技术,我们展示了使用MaxPeak Premier SEC 250 Å蛋白分析专用柱来提高Cas9的SEC-UV回收率,并开始研究不同化学计量比的核糖核蛋白(RNP)复合物的形成。另外,使用挥发性乙酸铵流动相通过直接的非变性质谱分析观察到约160 kDa的质量数,对应于游离链球菌Cas9蛋白的完整质量。对这些方法有初步认识后,人们相信将SEC分析用于其他CRISPR分子会取得更多的成功,如更小的(约124 kDa)金黄色葡萄球菌衍生的Cas9同源物。
优势
- 由于改善了色谱柱表面的惰性,MaxPeak Premier SEC 250 Å蛋白分析专用柱可实现优异的Cas9回收率
- Cas9、gRNA和RNP复合物的SEC-UV方法考量
- 初步了解Cas9的SEC-MS分析
简介
Francisco Mojica于1993年发明了成簇规律间隔短回文重复序列(CRISPR)1。 CRISPR是一种基因编辑技术,它需要两个组成部分:一个CRISPR关联(Cas)蛋白酶和一个向导RNA分子。近年来,CRISPR已成功应用于食品、植物、生物燃料生产中的基因编辑,现在又用于人类细胞的体外基因编辑和人类患者的体内基因编辑1。 最近,Jennifer Doudna和Emmanuelle Charpentier教授因对发现和使用CRISPR系统所做的贡献而获得了诺贝尔奖。在其中一些著名研究中,他们使用了大肠杆菌表达的重组Cas9(来自化脓链球菌)以及crispr RNA (crRNA)和体外转录反式激活crRNA(tracrRNA或trRNA)。他们证明由这些分子形成的复合物可以切割纯化的DNA,从而促进基因编辑(Jinek等人,2012)2。 此外,他们还表明,可以通过专门设计的crRNA编程靶位特异性,并且trRNA和crRNA可以融合成单个向导RNA (sgRNA)。sgRNA的概念现在已经广泛应用于基因组编辑1。
改变活细胞的基因组并不是一个新概念,在八十年代末就已经发生了革命性的变化,但被认为相对低效。基因组编辑技术,如设计者锌指(ZF)1–4、转录激活因子类效应物(TALE)4–6和归巢内切核酸酶5,都是基因编辑技术的重大突破。CRISPR技术已被证明在精确的DNA位点上执行双链DNA断裂更精准,并且可扩展、可负担且易于设计3-4。目前,治疗应用正在兴起。美国食品药品监督管理局(FDA)授予了Editas Medicine的EDIT-301罕见儿科疾病认定,这是一种用于治疗镰状细胞病(SCD)和输血依赖性β-地中海贫血(TDT)的CRISPR基因编辑研究药物。另外,目前福泰制药的CTX001正在进行III期临床试验,这是一种用于治疗β-地中海贫血的患者源CRISPR-Cas9编辑干细胞疗法5-6。
在自然和人工CRISPR/Cas系统中,CRISPR关联蛋白9 (Cas9)负责定位和切割靶DNA。本文中的II型Cas9蛋白是化脓链球菌中与CRISPR适应性免疫系统关联的双RNA引导DNA内切酶2,6。 这个约160 kDa的Cas9蛋白与crRNA和trRNA或单个向导RNA结合生成用于基因编辑的RNP复合物,可以进行特定的DNA切割。分子间的相互作用和设计对于用于治疗性基因编辑的RNP非常重要。如果没有仔细的设计和研究,可能会在细胞基因组中未指定的位点上发生脱靶切割。SEC技术已被证明是研究蛋白质及其复合物生物物理特性的一种有效的技术。蛋白质SEC的主要限制之一是静电和疏水次级相互作用,有些蛋白质特别容易出现这些问题。Cas9蛋白被认为难以使用SEC进行分析,原因可以通过其晶体结构的分析假设来解释(图1)。Shenal和同事观察到化脓链球菌Cas9的晶体结构中,有一个高度碱性的裂沟,以促进与其互补核酸的结合。Lys和Arg残基用蓝色突出显示,以说明裂沟的位置7。 尽管如此,链球菌Cas9也呈现酸性结构域。该基序的电负性可以解释为什么有报告称难以对这种蛋白质进行色谱分析,与Asp和Glu残基集中区域相对应的强电负性预计会对金属色谱柱硬件表现出亲和力,产生吸附损失。
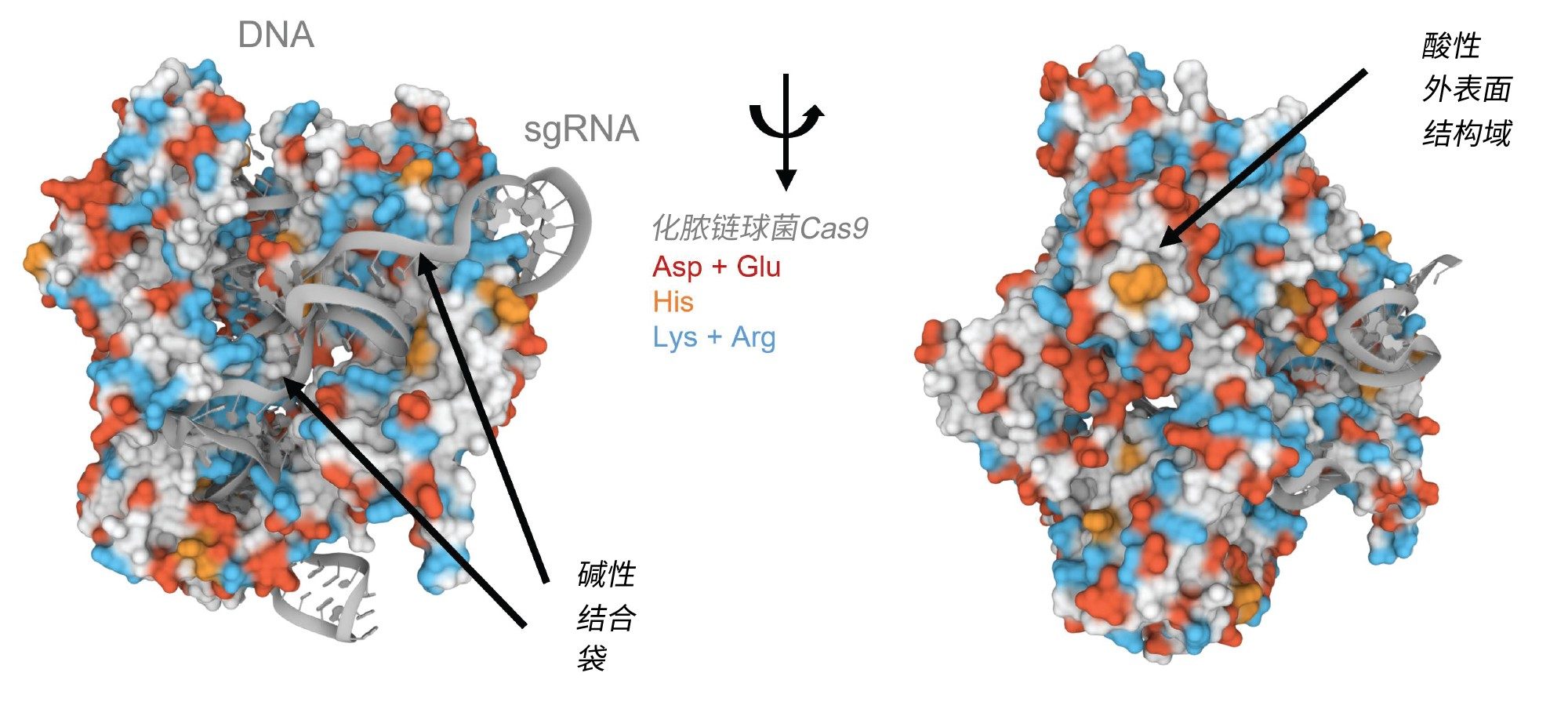 图1.化脓链球菌Cas9与向导RNA和靶DNA分子复合物的晶体结构。蛋白质残基显示在表面,核酸显示为底图。天冬氨酸和谷氨酸残基以红色突出显示,组氨酸残基以橙色突出显示,赖氨酸和精氨酸残基以蓝色突出显示。使用Mol*和RCSB PDB ID 4OO8创建图像。(D. Sehnal, S. Bittrich, M. Deshpande, R. Svobodová, K. Berka, V. Bazgier, S. Velankar, S.K. Burley, J. koka, A.S. Rose (2021) Mol* Viewer: modern web app for 3D visualization and analysis of large biomolecular structures. Nucleic Acids Research. doi:10.1093/nar/gkab31.7。)
图1.化脓链球菌Cas9与向导RNA和靶DNA分子复合物的晶体结构。蛋白质残基显示在表面,核酸显示为底图。天冬氨酸和谷氨酸残基以红色突出显示,组氨酸残基以橙色突出显示,赖氨酸和精氨酸残基以蓝色突出显示。使用Mol*和RCSB PDB ID 4OO8创建图像。(D. Sehnal, S. Bittrich, M. Deshpande, R. Svobodová, K. Berka, V. Bazgier, S. Velankar, S.K. Burley, J. koka, A.S. Rose (2021) Mol* Viewer: modern web app for 3D visualization and analysis of large biomolecular structures. Nucleic Acids Research. doi:10.1093/nar/gkab31.7。)
本应用纪要介绍简单的SEC方法,可用于Cas蛋白、CRISPR RNA及其形成的RNP复合物的表征。本研究应用了新开发的基于250 Å BEH™颗粒和羟基PEO键合的SEC填料,以及用亲水性MaxPeak高性能表面(h-HPS)改良的硬件。与传统的不锈钢色谱柱硬件相比,h-HPS表面硬件表现出对多种Cas9分子优异的回收率。h-HPS硬件显著提高了Cas9核酸酶的回收率,这预示着持续的方法开发和SEC CRISPR分子表征的应用(从药物设计到配方和稳定性研究)将会有更多机会。在一个案例中,已成功将基于磷酸盐缓冲液的流动相用于研究单个进样以及Cas9与RNA的比率混合物。已经观察到基于平衡的RNP复合物形成的迹象。另外,利用挥发性乙酸铵流动相和QTof质谱对约160 kDa的质量进行了直接的非变性质谱分析(MS)观察,该质量对应于游离链球菌Cas9蛋白的完整质量。对这些方法有了初步的认识后,人们相信将SEC分析用于CRISPR分子将会取得更多的成功,无论是将SEC用于多维分离、应用于非变性质谱分析,还是使用这些工具对更小的、更易处理的金黄色葡萄球菌衍生的Cas9同源物(~124 kDa)进行额外的研究。
实验
样品描述
Cas9核酸酶蛋白NLS购自Dharmacon/Horizon,含有化脓链球菌Cas9 (Csn1)基因(P/N: CAS12206),浓度为10 µg/µL,用milli-Q水稀释至5 µg/µL。Milli-Q水购自Millipore。通过向1升Milli-Q水中加入2包购自Sigma的PBS (P/N: P3813),制备pH 7.4的20 mM磷酸盐、276 mM NaCl、5.4 mM KCl(2x PBS缓冲液),并在使用前通过0.2 µm无菌尼龙过滤器过滤(Thermo Scientific,P/N: 1630020)。5倍稀释IonHance™ CX-MS浓缩液B制备320 mM乙酸铵,pH 8.5,溶于4% ACN。
RNP复合物形成
将购自Dharmacon/Horizon的sgRNA,即Edit-R Synthetic sgRNA Non-targeting Control #1 (P/N: U-009501-01-05)在60 °C下培养10分钟,再冷却至室温15分钟,从而形成RNP复合物。将加热后的sgRNA加入Cas9核酸酶蛋白NLS分别制备1:1和3:1的RNP复合混合物,然后轻微地旋涡混合。将混合物在8000 rpm下离心30秒,然后转移到沃特世聚丙烯材质12 x 32 mm螺口样品瓶,容积300 µL (P/N: 186002639)。
液相色谱条件
|
液相色谱系统: |
ACQUITY™ UPLC™ H-Class Bio |
|
检测条件: |
ACQUITY UPLC TUV检测器(钛合金流通池,5 mm,1500 nL) |
|
波长: |
280 nm(Cas9样品)和260 nm(RNP形成) |
|
数据采集: |
Empower™ Pro 3 Feature Release 3 |
|
样品瓶: |
最大回收样品瓶和瓶盖(沃特世P/N: 186000327C)和沃特世300 μL聚丙烯螺纹口样品瓶(沃特世P/N: 186004112) |
|
色谱柱: |
XBridge™ Premier SEC 250 Å, 2.5 µm, 4.6 x 150 mm蛋白分析专用柱 ACQUITY Premier SEC 250 Å, 1.7 µm, 4.6 x 300 mm蛋白分析专用柱 |
|
柱温: |
35 °C |
|
样品温度: |
8 °C |
|
样品管理器清洗液: |
18.2 MΩ水 |
|
密封清洗液: |
10% HPLC级甲醇/90% 18.2 MΩ水(v/v) |
|
进样体积: |
gRNA(2 µL,5 µg/µL);RNP形成(2 µL,1:1和3:1复合物) |
|
流速: |
0.2 mL/min |
|
流动相A: |
2X PBS:磷酸盐缓冲液(20 mM磷酸盐、276 mM NaCl、5.4 mM KCl,pH 7.4) |
|
样品: |
Cas9核酸酶蛋白NLS |
|
梯度: |
等度 |
SEC-MS
|
液相色谱系统: |
ACQUITY Premier二元溶剂管理器 |
|
检测条件: |
ACQUITY UPLC TUV检测器(钛合金流通池,5 mm,1500 nL) |
|
波长: |
280 nm |
|
数据采集 |
MassLynx™ 4.1版 |
|
样品瓶: |
最大回收样品瓶和瓶盖(沃特世P/N: 186000327C)和沃特世300 μL聚丙烯螺纹口样品瓶(沃特世P/N: 186004112) |
|
色谱柱: |
ACQUITY Premier SEC 250 Å, 1.7 µm, 4.6 x 150 mm蛋白分析专用柱 |
|
柱温: |
30 °C |
|
样品温度: |
8 °C |
|
密封清洗液和FTN SM清洗液: |
10% HPLC级甲醇/90% 18.2 MΩ水(v/v) |
|
进样体积: |
SpCas9 (10 µL) |
|
流速: |
0.1 mL/min |
|
流动相A: |
5倍稀释IonHance CX-MS浓缩液B制备320 mM乙酸铵,pH 8.5,溶于4% ACN |
|
样品: |
Cas9核酸酶蛋白NLS |
|
质谱系统: |
Xevo™ G2-XS QToF |
|
四极杆设置: |
1000/2000/3940 25%/25%/25%/25% |
|
采集窗口: |
2000~7000 m/z |
|
毛细管电压: |
3 kV |
|
扫描速率: |
2 Hz |
|
离子源温度: |
100 °C |
|
脱溶剂气流速: |
600 L/h |
|
脱溶剂气温度: |
450 °C |
|
锥孔气流速: |
50 L/h |
结果与讨论
之前的研究中已经探索了新型250 Å BEH HO-PEO键合颗粒和MaxPeak h-HPS硬件在SEC过程中减少静电和疏水次级相互作用的优势8。 在本研究中,我们展示了可以应用平台方法和基于2倍浓度磷酸盐缓冲液(20 mM磷酸盐、276 mM NaCl、5.4 mM KCl,pH 7.4;2 x PBS)的流动相。这为化脓链球菌Cas9的色谱研究提供了一个起点。首先通过实验比较了用SEC 250 Å蛋白分析专用柱填料和标准金属硬件与经h-HPS改良的硬件制备的SEC色谱柱的峰形和回收率。具体来说,我们比较了传统不锈钢色谱柱硬件和h-HPS色谱柱硬件的性能,色谱柱尺寸均为4.6 x 300 mm,使用相同的羟基封端BEH 250 Å, 2.5 µm填料。图2展示了得到的色谱图。正如预测的那样,典型的CRISPR Cas9蛋白上高度集中的静电结合位点会导致传统不锈钢硬件上产生吸附损失。
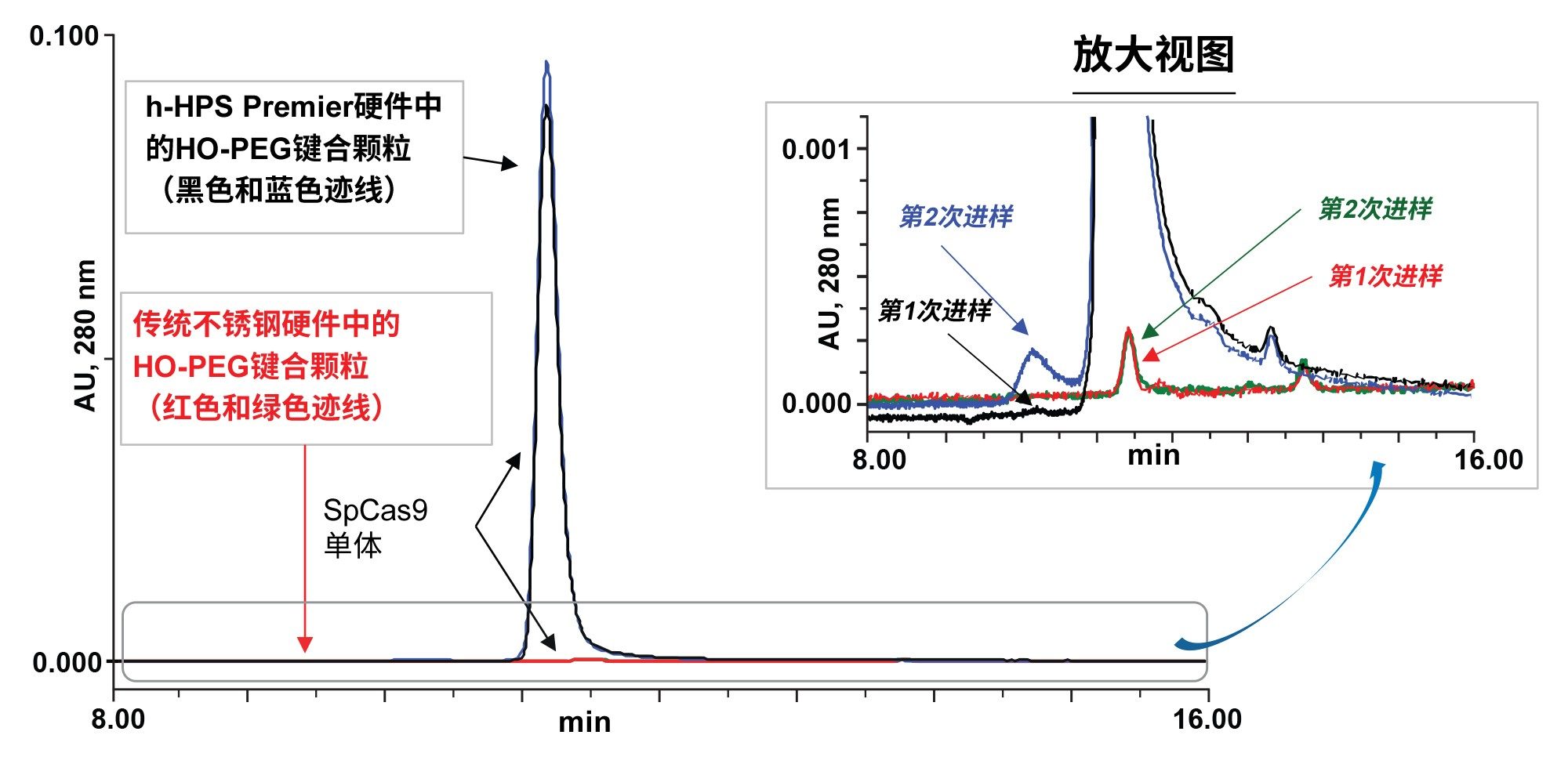 图2.SpCas9蛋白使用传统不锈钢色谱柱硬件和亲水性MaxPeak高性能表面色谱柱硬件的对比,使用250 Å BEH颗粒和高覆盖率羟基封端PEO键合以及2x PBS流动相。化脓链球菌Cas9中的SpCas9核酸酶蛋白在进样1和2中与传统不锈钢硬件(红色和绿色迹线)的静电相互作用分别强于h-HPS色谱柱硬件(黑色和蓝色迹线)中的静电相互作用。
图2.SpCas9蛋白使用传统不锈钢色谱柱硬件和亲水性MaxPeak高性能表面色谱柱硬件的对比,使用250 Å BEH颗粒和高覆盖率羟基封端PEO键合以及2x PBS流动相。化脓链球菌Cas9中的SpCas9核酸酶蛋白在进样1和2中与传统不锈钢硬件(红色和绿色迹线)的静电相互作用分别强于h-HPS色谱柱硬件(黑色和蓝色迹线)中的静电相互作用。
传统不锈钢和h-HPS硬件之间的性能差异巨大。在图2中,使用BEH-PEO颗粒的h-HPS色谱柱硬件显著降低了SpCas9的静电性能(图1)。使用传统的不锈钢色谱柱硬件几乎不可能回收SpCas9。有理由认为,SpCas9的静电域强烈吸附在色谱柱内的金属硬件表面。即使使用h-HPS硬件,也有细微的活化迹象,即与第一次进样相比,第二次进样的总回收率有所增加(蓝色迹线)。尽管如此,h-HPS色谱柱上第一次进样的SpCas9单体回收率依然非常高。SpCas9的稳定性效应尚未得到研究以充分了解聚集体形成的速度。
由于使用h-HPS硬件,XBridge™ Premier SEC 250 Å, 2.5 µm蛋白分析专用柱性能良好,因此将这种色谱柱技术应用于Cas9蛋白的额外研究,以及研究RNP复合物的形成和色谱行为。
RNP复合物形成
Cas9的构象变化尚未得到充分了解,但Jinek和同事推测蛋白质侧链与RNA碱基之间的空间相互作用或弱结合可能会导致这些变化10。 在RNP复合物的形成过程中,快速评估络合过程中和络合后化合物的稳定性至关重要。要通过SEC进行评估,我们使用ACQUITY Premier SEC 250 Å 1.7µm, 4.6 x 300 mm蛋白分析专用柱,并以易于制备的磷酸盐缓冲液作为流动相研究RNP的形成。下图3a-d显示了单个sgRNA、单个SpCas9和RNP复合物的SEC-UV迹线,这些复合物分别由sgRNA和SpCas9以1:1和3:1混合而成。
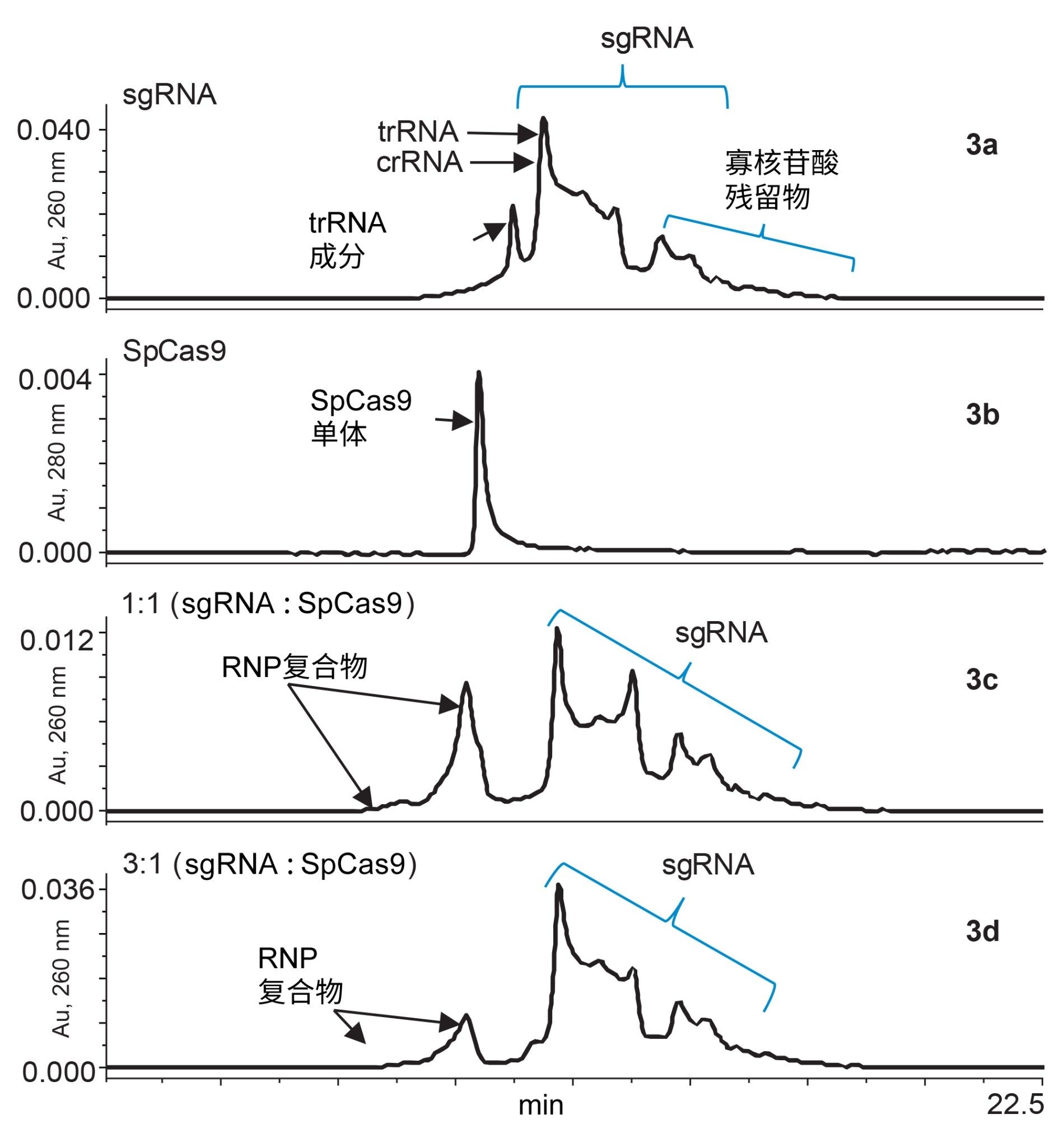 图3a-d.使用ACQUITY Premier SEC 250 Å 1.7µm 4.6 x 300 mm蛋白质分析专用柱获得的RNP复合物形成、单个向导RNA和Cas9蛋白的色谱图。图3a为在260 nm处获得的sgRNA的UV色谱图。图3b为在280 nm处获得的化脓链球菌Cas9的色谱图。图3c显示了sgRNA和Cas9以1:1混合时RNP复合物的形成。图3d显示了sgRNA和Cas9以3:1混合时RNP复合物的形成。
图3a-d.使用ACQUITY Premier SEC 250 Å 1.7µm 4.6 x 300 mm蛋白质分析专用柱获得的RNP复合物形成、单个向导RNA和Cas9蛋白的色谱图。图3a为在260 nm处获得的sgRNA的UV色谱图。图3b为在280 nm处获得的化脓链球菌Cas9的色谱图。图3c显示了sgRNA和Cas9以1:1混合时RNP复合物的形成。图3d显示了sgRNA和Cas9以3:1混合时RNP复合物的形成。
图3a为分子量约为32 kDa的编辑sgRNA的SEC-UV色谱图。该色谱图在UV 260 nm下测得,以更有选择性地监测核酸洗脱。同时,图3b为SpCas9的SEC-UV 280 nm色谱图,对应的单体峰在11.34分钟洗脱。图3c为sgRNA与SpCas9以1:1混合的结果。由于在9.00-11.09分钟之间出现了新的洗脱峰,证明RNP复合物已部分形成。这些RNP峰的洗脱移位可能是由于结合的蛋白质-RNA复合物的流体动力学半径增加。通过RNP复合物峰上的肩峰可以发现,在1:1比例下有一小部分SpCas9蛋白质未结合。图3d中,sgRNA的比例增加到3:1,增加后产生了大量游离sgRNA的信号。随着sgRNA过量,未结合的SpCas9肩峰也相应地减少。虽然还有很多有待了解的地方,但SEC-UV分析可以用于RNP络合的快速检查,并作为监测其稳定性的工具。通过研究用于形成RNP的sgRNA与Cas9蛋白的不同比例,也可以评估结合亲和力,进而用于药物设计实验。
挥发性流动相和SEC-MS的初步研究
为了增强对Cas蛋白及其复合物的表征,可以同时获取洗脱分析物的质谱信息。不仅使用了上述磷酸盐缓冲液流动相研究SpCas9的分离,还使用了可以与在线MS检测直接连接的乙酸铵溶液。在这类研究中,只能使用微量金属和MS认证的流动相组分,这非常重要,因为钠和钾加合物很容易对MS质量造成不利影响。为此,采用IonHance乙酸铵浓缩液,并进行了一系列不同稀释因子的研究实验。使用100 mM、200 mM、240 mM、280 mM、300 mM和400 mM乙酸铵对应的UV色谱图如图4所示。即使使用基于磷酸盐的缓冲液,SpCas9也会遇到一些问题,因此可以理解至少需要约300 mM的乙酸铵才能获得对称的单体峰和高回收率。虽然本研究中没有进行研究,但有理由相信其他pH滴定乙酸铵溶液可能同样或更好地适用于这种应用。
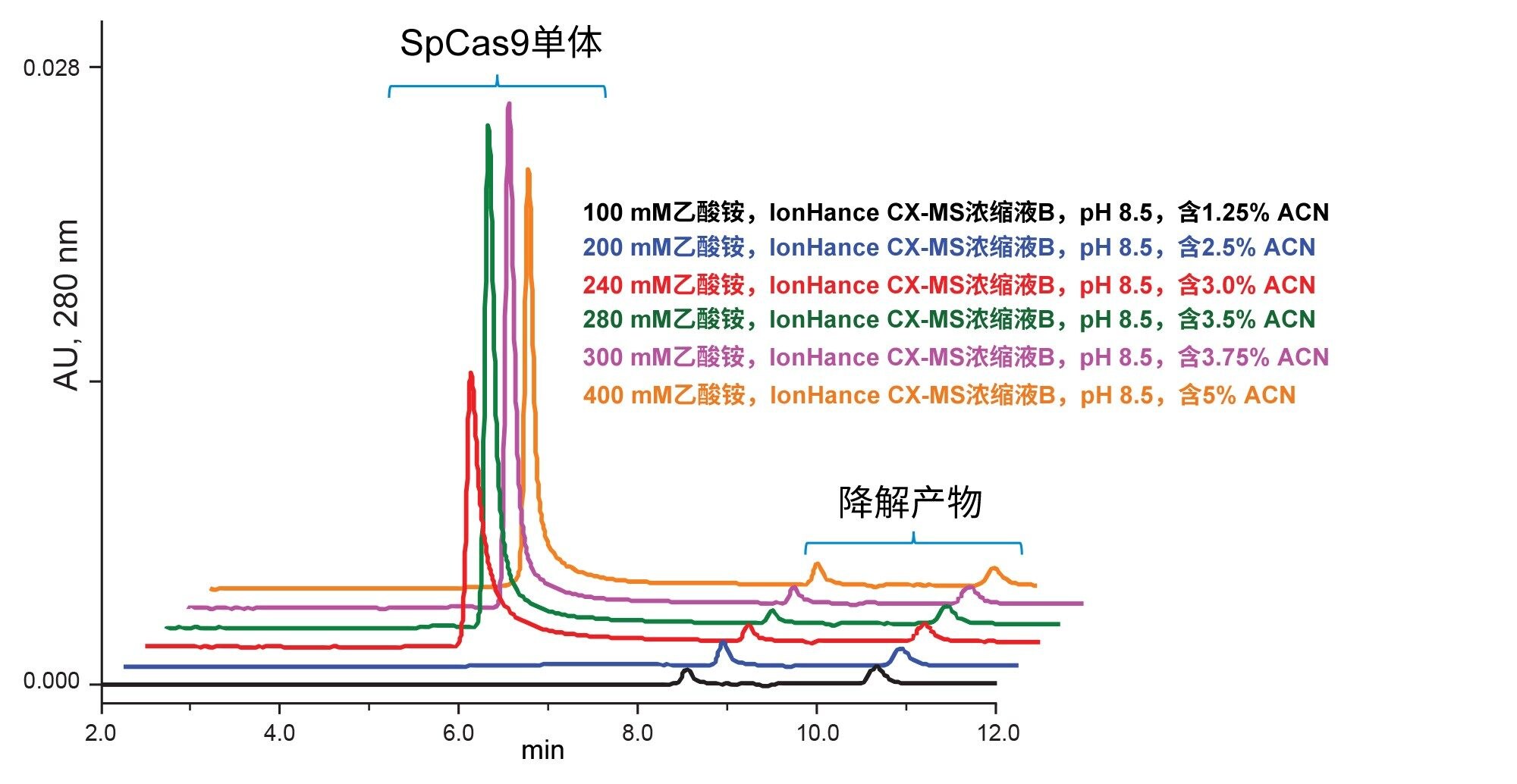 图4.使用IonHance浓缩液B缓冲液,稀释为pH 8.5的乙酸铵流动相,包括100 mM、200 mM、240 mM、280 mM、300 mM和400 mM,进行SpCas9分析。
图4.使用IonHance浓缩液B缓冲液,稀释为pH 8.5的乙酸铵流动相,包括100 mM、200 mM、240 mM、280 mM、300 mM和400 mM,进行SpCas9分析。
与非变性LC-MS通常应用的值相比,320 mM乙酸铵流动相相对较高。尽管如此,我们研究了QTof质谱的优化方法,以检测高m/z范围内的物质。调整了四极杆质量数过滤以使低m/z离子不稳定,而对2000 m/z及以上离子实现更有选择性的检测。流速为0.1 mL/min,以减少ESI离子源的脱溶剂负担,进而得到如图5A所示的UV和总离子流色谱图。实验中使用的320 mM乙酸铵流动相使单体SpCas9及其HMW物质都产生了良好的峰形。放大的色谱图(见插图)凸显了这种分离的质量。SpCas9是一种大分子蛋白,预计分子量为158 kDa (UniProt Q99ZW2)。它还被预测有多个镁离子强结合袋,并且可能对其进行转译后修饰。此外,本文研究的SpCas9结构是由Csn1基因与结合C端核定位信号(NLS)重组制备的,由此增加了一段碱性残基,总质量为1~3 kDa。图5B所示为由单体峰组成的汇总原始质谱图,以及MaxEnt1去卷积谱图。在原始质谱图中观察到异质性,对其进行适当地处理以测定分子量信息。从这个实验中去卷积得到的两个强度最高的质量数是161.5和161.9 kDa,都与预测的Csn1基因产物的平均质量和NLS C端延伸(重量在1至3 kDa之间)相一致。这些结果表明,基于MS的Cas9蛋白检测已经触手可得,并且额外的方法开发可以提高灵敏度和MS质量,提供检测Cas9蛋白与其向导RNA分子络合作用所需的分析能力。还应该注意的是,将SEC-MS分析用于更小的(约124 kDa)金黄色葡萄球菌Cas9同源物可以取得更多的成功。
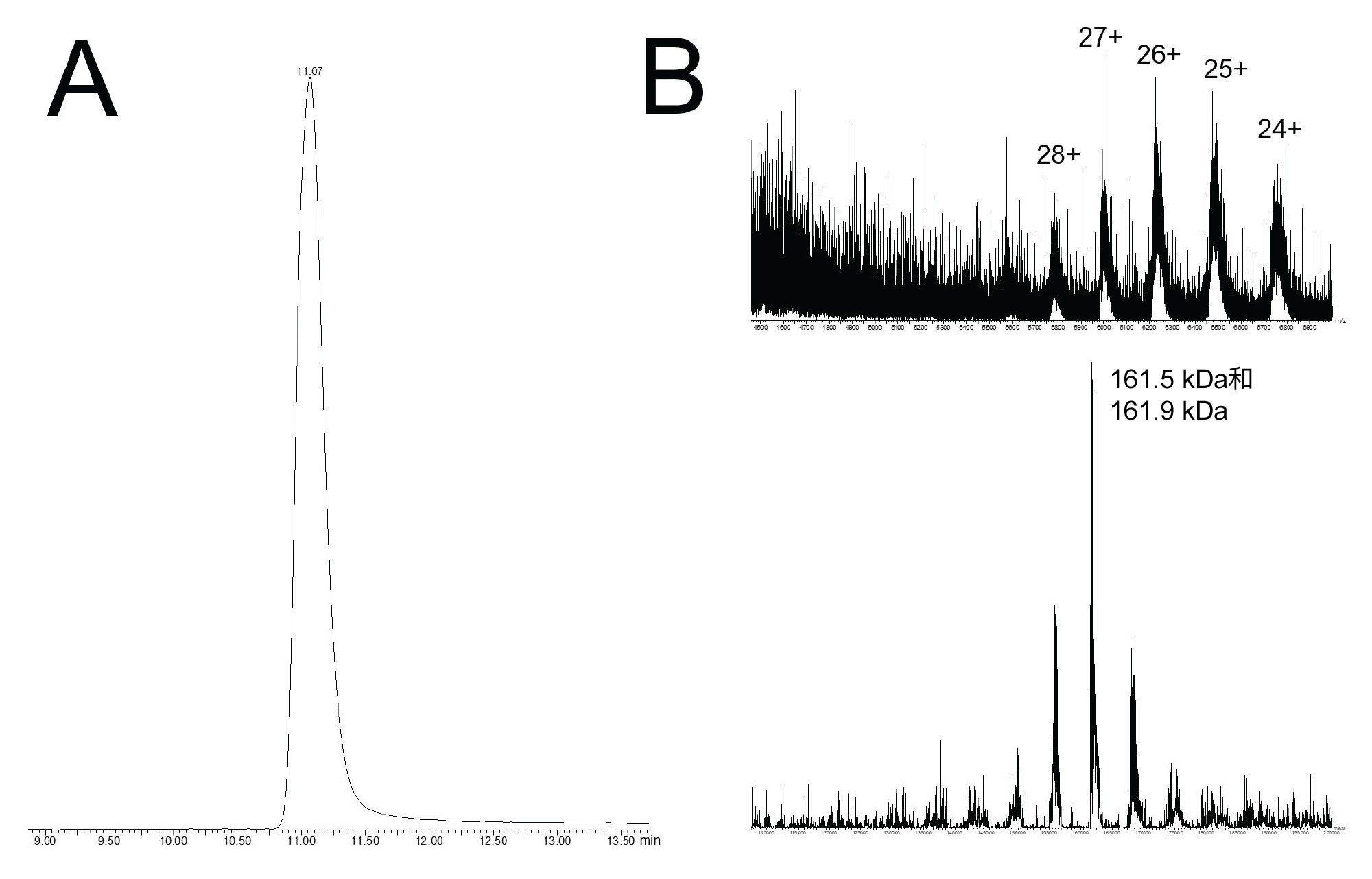 图5.使用ACQUITY Premier SEC 250 Å 1.7µm 4.6 x 150 mm蛋白分析专用柱和320 mM乙酸铵流动相(pH 8.5)的非变性SEC-MS。(A)由QTof ESI-MS所得的总离子流色谱图。(B)单体SpCas9物质对应的汇总原始质谱图及其对应的MaxEnt1去卷积谱图。
图5.使用ACQUITY Premier SEC 250 Å 1.7µm 4.6 x 150 mm蛋白分析专用柱和320 mM乙酸铵流动相(pH 8.5)的非变性SEC-MS。(A)由QTof ESI-MS所得的总离子流色谱图。(B)单体SpCas9物质对应的汇总原始质谱图及其对应的MaxEnt1去卷积谱图。
结论
在本应用纪要中,我们创建了一个简单的SEC-UV方法来分析CRISPR/Cas9蛋白及其RNP复合物。本研究使用了Waters ACQUITY Premier SEC 250 Å, 1.7µm蛋白分析专用柱和XBridge Premier SEC 250 Å, 2.5µm蛋白分析专用柱技术,这两种色谱柱允许使用简单的PBS基流动相。先前的研究表明,这些新型色谱柱的表面技术显著减少了蛋白质与色谱柱的非特异性相互作用10。 本研究发现,使用新的基于h-HPS的色谱柱技术显著减少了化脓链球菌Cas9蛋白在常规硬件色谱柱上的静电次级相互作用。这提高了SpCas9分析的准确度,并提供了开发在线SEC-MS方法的机会,该方法使用经MS认证的由IonHance乙酸铵组成的流动相缓冲液。这两种SEC方法都显示出对CRISPR/Cas9及其对应核酸,以及RNP复合物形成进行研究的潜力。
参考资料
- Lander Eric S (2016) The Heroes of CRISPR.Cell 164(1):18–28.doi.org/10.1016/j.cell.2015.12.041.
- Jinek M, Chylinski K, Fonfara I, Hauer M, Doudna JA, Charpentier E. A Programmable Dual-RNA-Guided DNA Endonuclease in Adaptive Bacterial Immunity. Science. 2012;337(6096):816–821.doi:10.1126/science.1225829.
- Le Cong, F. Ann Ran, David Cox, Shuailiang Lin, Robert Barretto, Naomi Habib, Patrick D. Hsu, Xuebing Wu, Wenyan Jiang, Luciano A. Marraffini, and Feng Zhang.Multiplex Genome Engineering Using CRISPR/Cas Systems.Science.2013 February 15; 339(6121): 819–823.doi:10.1126/science.1231143.
- Prashant Mali, Luhan Yang, Kevin M. Esvelt, John Aach, Marc Guell, James E. DiCarlo, Julie E. Norville, and George M. Church.RNA-Guided Human Genome Engineering via Cas9.Science.2013 February 15; 339(6121): 823-826.doi:10.1126/science.1232033.
- CRISPR Medicine News.News: Gene-Editing Clinical Trials for Beta Thalassemia - CRISPR Medicine (crisprmedicinenews.com).
- Deltcheva E, Chylinski K, Sharma CM, Gonzales K, Chao Y, Pirzada ZA, Eckert MR, Vogel J, Charpentier E. CRISPR RNA Maturation by trans-Encoded Small RNA and Host Factor RNase III.Nature. March 2011; 471(7340): 602–607. doi:10.1038/nature09886.
- David Sehnal, Sebastian Bittrich, Mandar Deshpande, Radka Svobodová, Karel Berka, Václav Bazgier, Sameer Velankar, Stephen K Burley, Jaroslav Koča, Alexander S Rose: Mol* Viewer: Modern Web App for 3D Visualization and Analysis of Large Biomolecular Structures, Nucleic Acids Research, 2021; 49(W1):431-437.doi:10.1093/nar/gkab31.
- Lavelay Kizekai, Stephen J. Shiner, Matthew A. Lauber.Waters ACQUITY和XBridge Premier SEC 250 Å蛋白分析专用柱:惰性SEC色谱柱设计的新基准.沃特世应用纪要, 720007493ZH, 2022.
- Jinek M., Jiang F., Taylor D. W., Sternberg S. H., Kaya E., Ma E., Anders C., Hauer M., Zhou K., Lin S., Kaplan M., Iavarone A. T., Charpentier E., Nogales E., Doudna J. A., Structures of Cas9 Endonucleases Reveal RNA-Mediated Conformational Activation. Science. 2014; 343(6176).doi/10.1126/science.1247997.
- Stephan M. Koza, Hua Yang, and Ying Qing Yu.在生理pH值和离子强度下通过现代体积排阻色谱法分离生物类似药抗体.沃特世应用纪要, 720007484ZH, 2022.
720007727ZH,2022年9月