使用集成式UPLC-MS和信息学工作流程进行RNA酶解产物图谱分析
摘要
LC-MS分析具有高灵敏度、高专属性并且能够检测多种寡核苷酸修饰,是单向导RNA (sgRNA)分析不可或缺的工具。本应用纪要重点介绍了几项新型信息学工具,这些工具用于快速处理sgRNA酶解后采集的LC-MS数据集。新的工作流程采用了包括waters_connect™ MAP Sequence应用程序在内的数据处理软件。使用mRNA Cleaver MicroApp模拟酶解的寡核苷酸产物,使用MAP Sequence应用程序匹配实验数据,并使用Coverage Viewer MicroApp查看结果。本文所述工作流程重点介绍使用一系列RNase T1、T2(RapiZyme™ MC1和Cusativin)和hRNase4核糖核酸酶的样品前处理步骤,以及LC-MS分析和数据处理流程。多种酶的特异性提供了独特的酶切位点,再结合新的信息学工具(利用准确质量数测量),可以快速、高效地确认sgRNA序列,从而提高药物开发效率,并加快RNA类产品面向临床或客户的投放进程。
优势
- 符合法规要求的全新信息学工具简化了使用UPLC-MS的RNA产物图谱分析过程
- 与常规的RNase T1酶解物相比,RNase T2提供了独特的酶切位点,酶解产物重叠的可能性更高
- 将一组酶解结果相结合,可提高对质量指纹图谱分析方法准确度的整体信心,有机会实现更高或完全的序列覆盖率
简介
2012年发表的原始CRISPR论文中首次描述了单向导RNA (sgRNA),当时将裂解双链DNA所需的两个RNA分子(tracRNA - Cas9酶支架和crRNA - 参与DNA靶标识别)融合成一个RNA结构1。sgRNA是CRISPR-Cas9基因编辑系统的关键组成部分1-2。 sgRNA分子引导Cas9核酸酶在DNA中产生双链断裂,打开了基因编辑的大门。CRISPR技术的发现彻底改变了基因编辑应用领域,并因此荣获2020年诺贝尔化学奖。CRISPR的主要应用之一是在2020年全球大流行疫情期间开发简单、快速的COVID-19检测方法,此外,CRISPR技术在研究和个性化医疗中也有许多其他应用,包括在治疗遗传性疾病、癌症和传染病中的应用³。sgRNA通常采用固相寡核苷酸合成方法产生,其分析表征包括完整分子量确证和完整序列确证4,5。
目前的sgRNA酶解产物图谱分析工作流程需要手动分析数据,费时费力。RNA酶解策略尚未经过优化,通常会产生大量模糊的寡核苷酸酶解产物,这些产物对于RNA分子序列的确认作用甚微。在2023年发布的应用纪要中,我们讨论了UPLC-MS和信息学工作流程,其开发目的是在完成RNase T1酶解后自动分析sgRNA和mRNA的序列图谱6。 本应用纪要详细介绍了一种修订后的工作流程,利用多种酶独立反应,通过累积明确的酶解产物分配来提高序列覆盖率。本文报告了一系列酶解得到的酶解产物图谱分析结果,包括RNase T1、T2(RapiZyme MC1和Cusativin)和hRNase4。该工作流程的另一项重大改进在于引入了新的waters_connect应用程序(MAP Sequence),用于快速(不到3分钟)、高效地自动分配RNA酶解产物图谱。
实验
试剂和样品前处理
二异丙基乙胺(DPA,纯度99%,产品目录号D214752–500ML)和1,1,1,3,3,3-六氟异丙醇(HFIP,纯度99%,产品目录号105228-100G)购自Millipore Sigma(美国密苏里州圣路易斯)。甲醇(LC-MS级,产品目录号34966–1L)购自Honeywell(美国卡罗来纳州夏洛特)。HPLC级I级去离子(DI)水使用Milli-Q系统(Millipore,美国马萨诸塞州贝德福德)净化。流动相每日现配现用。用于sgRNA/mRNA酶解的无核酸酶超纯水(产品目录号J71786.AE)购自Thermo Fisher Scientific(美国马萨诸塞州沃尔瑟姆)。
从Integrated DNA Technologies(美国艾奥瓦州科尔维尔)购得10 nmol的100 mer sgRNA寡核苷酸编码HPRT1酶(次黄嘌呤磷酸核糖基转移酶1)。sgRNA寡核苷酸的序列为:5’-A*A*A* UCC UCA GCA UAA UGA UUG UUU UAG AGC UAG AAA UAG CAA GUU AAA AUA AGG CUA GUC CGU UAU CAA CUU GAA AAA GUG GCA CCG AGU CGG UGC U*U*U* U-3’。RNA寡核苷酸的前三个5'-端核苷酸(A*A*A*)以及最后三个3'-端核苷酸(U*U*U*)上含有2'-OMe修饰(星号表示这六个核苷酸均被硫代磷酸化)。用去离子水制备sgRNA寡核苷酸的储备液,浓度为5 µM。
真菌源性不含动物源组分的纯化RNase T1(产品目录号IFGRNASET1AFLY 500KU)购自Innovative Research(美国密歇根州诺维),将冻干酶溶于5 mL 100 mM碳酸氢铵(产品目录号5.33005–50G,Millipore Sigma)中,制备100单位/μL的溶液。hRNase4酶(产品目录号M1284S)购自New England Biolabs(NEB,马萨诸塞州伊普斯威奇)。RapiZyme MC1(P/N:186011190,10000单位/管)和RapiZyme Cusativin(P/N:186011192,10000单位/管)是沃特世公司最近推出的两款新型RNA酶7。
使用RNase T1酶解sgRNA时,将2 μL 5 µM sgRNA与28 μL无核酸酶水和10 μLRNase T1混合,在37 oC下酶解15 min。在QuanRecovery MaxPeak 300 µL样品瓶中制备酶解混合物,随后立即以5 μL进样体积对酶解物进行LC-MS分析。
使用hRNase4酶解sgRNA时,将2 μL 5 µM sgRNA的3M尿素溶液在90 oC下热变性5分钟,然后快速冷却至25 oC。将变性的sgRNA与4 μL酶缓冲液(10倍浓度)、33 μL无核酸酶水和1 μL hRNase4酶(50单位/μL)混合,在37 oC下酶解60 min。用1 μL NEB核糖核酸酶抑制剂(NEB产品目录号M0314S)终止hRNase4酶解,然后在室温下温育10 min。
RapiZyme MC1和Cusativin的酶解方案非常相似。使用RapiZyme MC1时,将sgRNA(10 μL,2-5 µM溶液)置于含有200 mM乙酸铵的缓冲液(pH 8.0)中,在90 oC下处理2 min使其变性。使用RapiZyme Cusativin时,将sgRNA(10 μL,2–5 µM溶液)置于含有200 mM乙酸铵的缓冲液(pH 9.0)中,在90 oC下处理2 min使其变性。两种样品均置于冰上冷却,然后置于微量离心机中旋转以收集样品液滴。加入50单位的酶(1 μL RapiZyme MC1或Cusativin)和8 μL无核酸酶水,使最终体积约20 μL,然后在Eppendorf恒温混合器中在37 oC下将sgRNA酶解60 min。将溶液暴露于70 ºC下15 min使酶失活,终止酶解。随后立即以5 μL进样体积对酶解物进行LC-MS分析。
所有数据集均使用UNIFI™科学信息系统应用程序3.6.0.21版采集,随后使用waters_connect信息学平台内的MAP Sequence应用程序在mRNA Cleaver和Coverage Viewer两款MicroApp的协助下处理数据。
液相色谱条件
|
LC-MS系统: |
Xevo™ G3 QTof LC-MS搭配ACQUITY™ Premier系统 |
|
色谱柱: |
ACQUITY™ Premier BEH C18 FIT寡核苷酸分析专用柱, 130 Å, 1.7 µm, 2.1 x 150 mm(P/N:186009487) |
|
柱温: |
60 oC |
|
流速: |
300 µL/min |
|
流动相: |
溶剂A:10 mM DPA(二异丙基乙胺),40 mM HFIP(1,1,1,3,3,3-六氟异丙醇),溶于去离子水中,pH 8.5 溶剂B:10 mM DPA,40 mM HFIP,溶于50%甲醇中 |
|
样品温度: |
8 oC |
|
样品瓶: |
QuanRecovery MaxPeak HPS样品瓶(P/N:186009186) |
|
进样体积: |
5 µL |
|
清洗溶剂: |
灌注溶剂:50%甲醇 样品管理器清洗溶剂:50%甲醇 密封清洗液:含20%乙腈的去离子水溶液 |
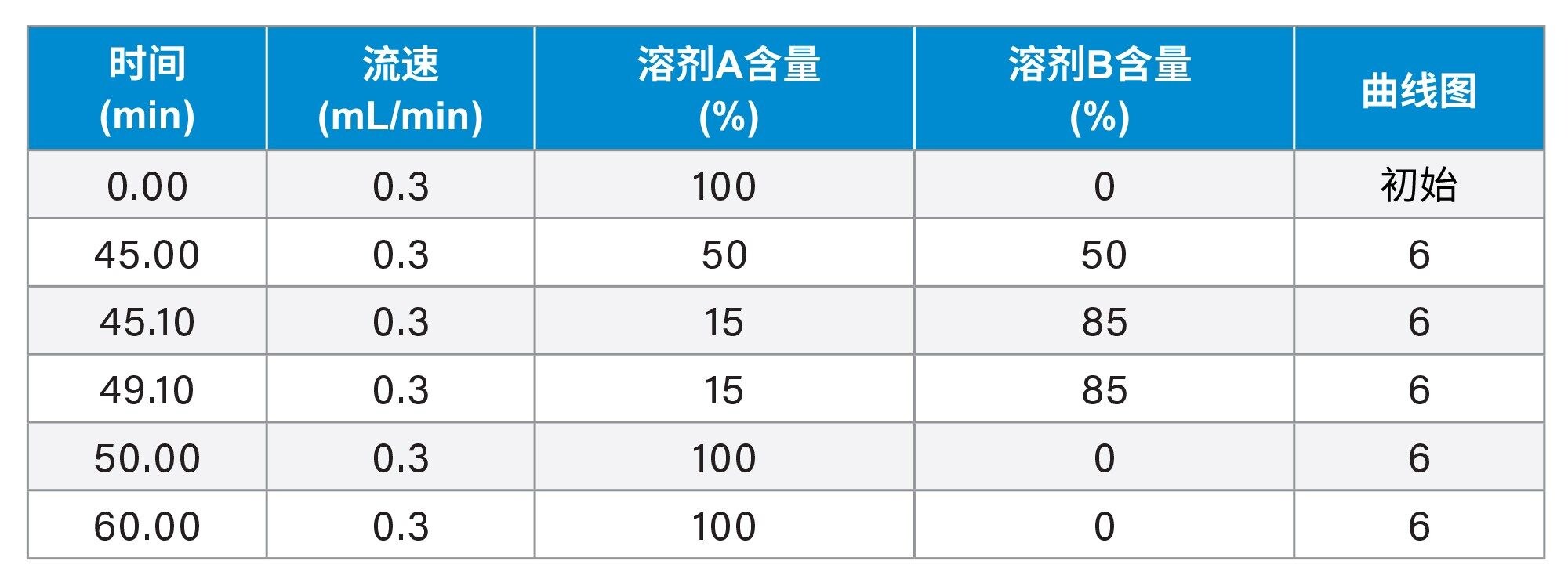
梯度表
质谱条件
|
质谱系统: |
Xevo™ G3 QTof质谱仪 |
|
电离模式: |
ESI(-) |
|
采集模式: |
MSE |
|
采集速率: |
1 Hz |
|
毛细管电压: |
2.5 kV |
|
锥孔电压: |
40 V |
|
离子源补偿: |
60 V |
|
离子源温度: |
120 oC |
|
脱溶剂气温度: |
550 oC |
|
锥孔气流速: |
50 L/h |
|
脱溶剂气流速: |
600 L/h |
|
TOF质量范围: |
340-4000(MSE采集) |
|
低能量CE: |
6 V |
|
高能量CE梯度: |
25~50 V |
|
实时校正标准液: |
50 pg/μL亮氨酸脑啡肽 |
|
数据采集: |
waters_connect 3.6.0.21 |
|
数据处理: |
waters_connect 3.6.0.21 |
|
数据处理: |
mRNA Cleaver 2.0 MAP Sequence v 1.0 Coverage Viewer v 2.0 |
结果与讨论
生物治疗药物研究机构采用LC-MS方法来确认RNA序列的情况越来越普遍。色谱重现性和分离度的改善,高质量分辨率MS的可用性增加,以及MS系统灵敏度和准确度的提升,都有助于在序列确认和碱基修饰的检测中获得可靠的结果。最重要的是,新型寡核苷酸信息学工具让RNA图谱分析中艰巨的数据分析任务变得更加轻松。
工作流程特点
图1突出显示了sgRNA经各种核糖核酸酶酶解后获得的LC-MSE数据集的处理工作流程。该工作流程的第一步是使用mRNA Cleaver MicroApp生成模拟酶解的寡核苷酸产物。第二步是使用waters_connect MAP Sequence应用程序自动将预测的寡核苷酸酶解产物分配给LC-MS数据。在图1所示工作流程的最后一步,使用Coverage Viewer MicroApp汇总经过多种酶酶解的sgRNA获得的序列覆盖率。
图2显示了mRNA Cleaver MicroApp的GUI(图形用户界面)示例,以及RNase T1酶解经修饰sgRNA所用的设置。mRNA Cleaver MicroApp中默认提供其他几种酶,包括MazF、RNase A、hRNase4、Colicin E5,以及沃特世公司最近推出的两种酶:RapiZyme™ MC1和RapiZyme™ Cusativin。
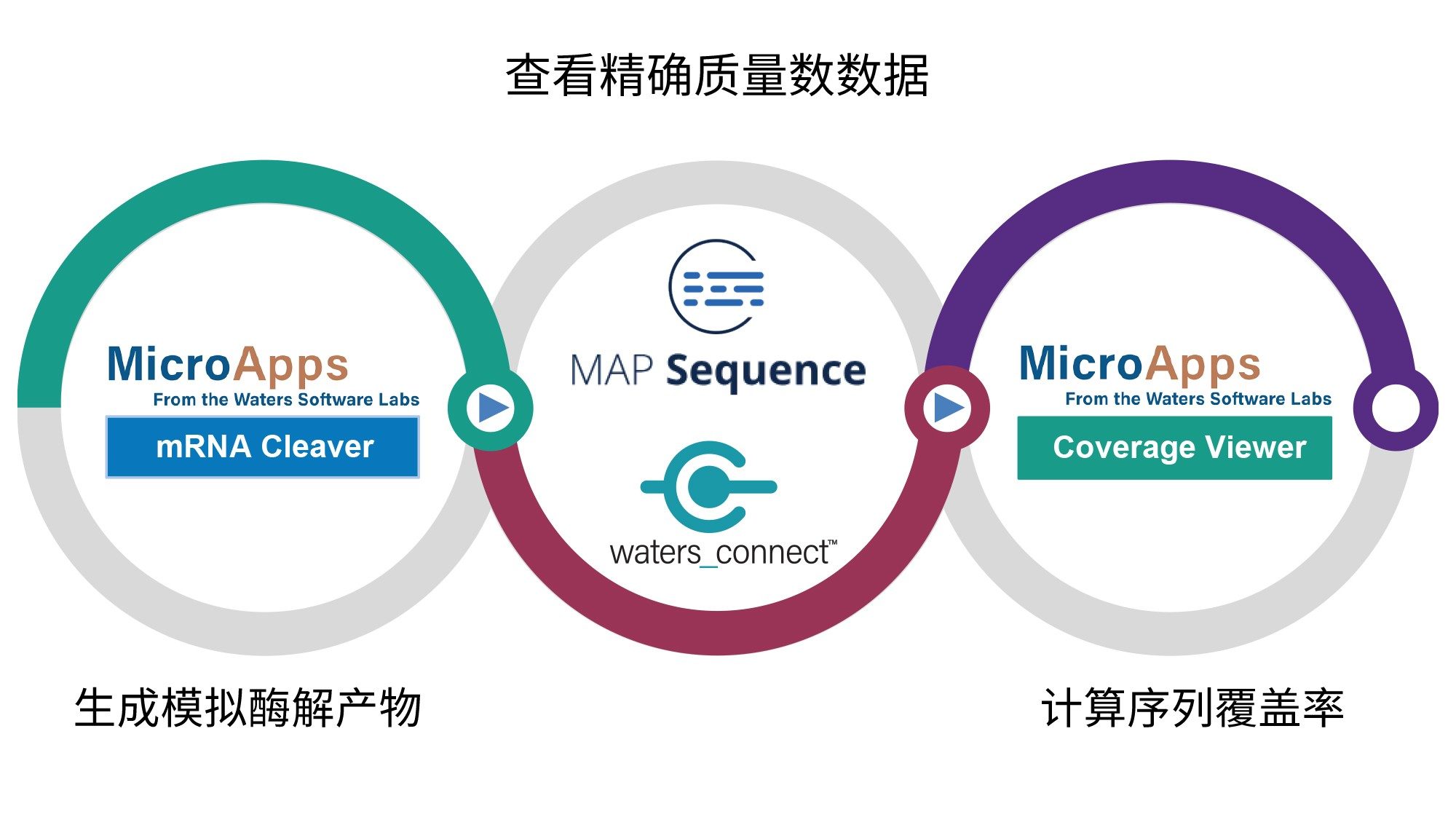 图1.使用mRNA Cleaver MicroApp、MAP Sequence应用程序和Coverage Viewer MicroApp对sgRNA/mRNA酶解物进行LC-MSE处理的工作流程简单示意图。
图1.使用mRNA Cleaver MicroApp、MAP Sequence应用程序和Coverage Viewer MicroApp对sgRNA/mRNA酶解物进行LC-MSE处理的工作流程简单示意图。
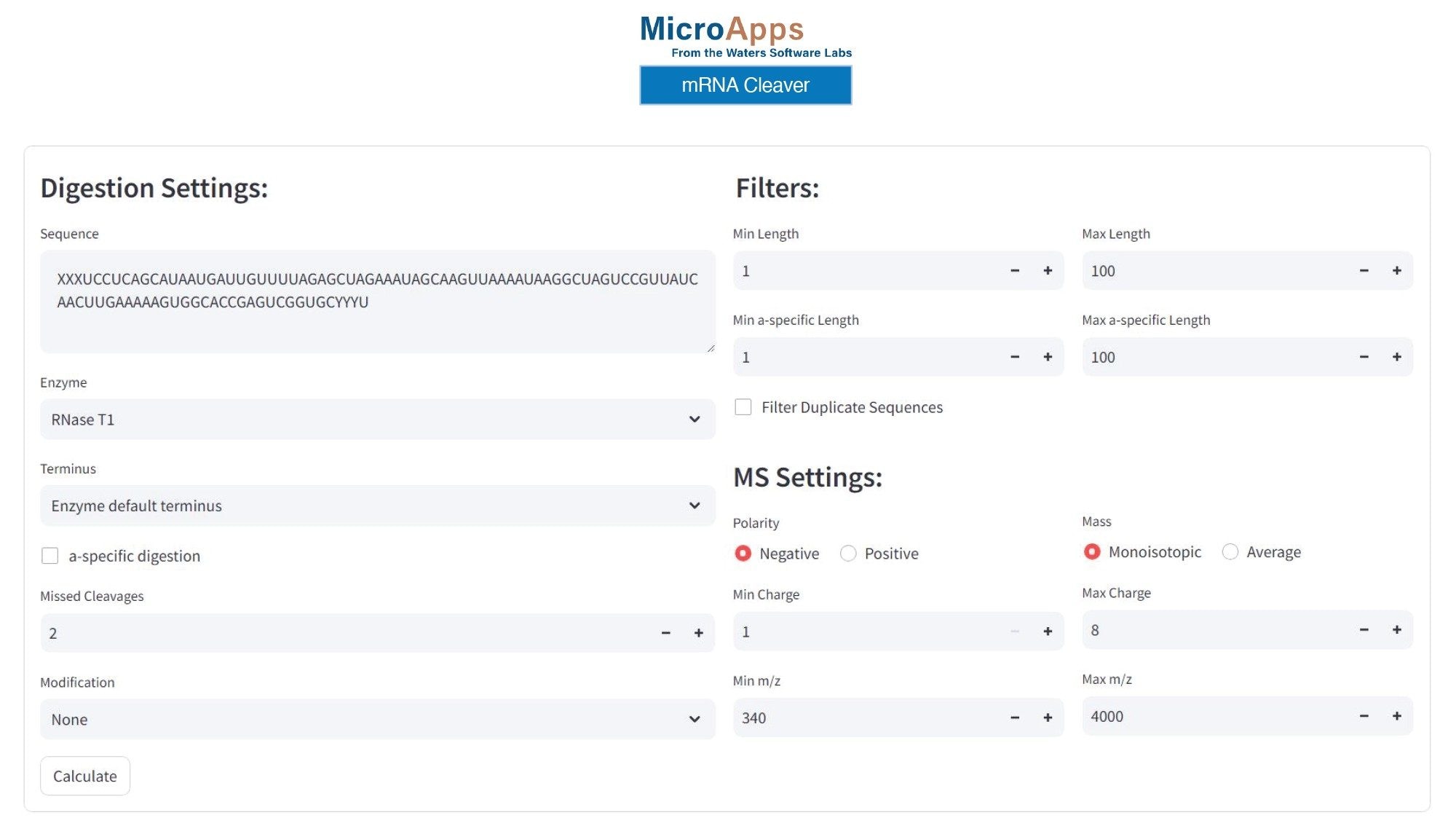 图2.mRNA Cleaver对RNase T1酶切经修饰100 mer sgRNA所得的寡核苷酸产物进行模拟预测所用的设置。残基X表示2'-OMe硫代磷酸化腺苷,残基Y表示2'-OMe硫代磷酸化尿苷。
图2.mRNA Cleaver对RNase T1酶切经修饰100 mer sgRNA所得的寡核苷酸产物进行模拟预测所用的设置。残基X表示2'-OMe硫代磷酸化腺苷,残基Y表示2'-OMe硫代磷酸化尿苷。
正如实验部分所述,RNA寡核苷酸的前三个5'-端核苷酸(A*A*A*,在图2所示的RNA序列中用字母XXX表示)含有2'-OMe修饰和硫代磷酸化连接子。此外,图2所示序列中最后三个3'-端核苷酸(U*U*U*,用字母YYY表示)被硫代磷酸化并含有2'-OMe官能团修饰。使用图2所示设置预测的模拟酶解产物,假设RNase T1酶存在至多两个漏切,生成的寡核苷酸产物在3'-位含有线性磷酸酯,并被视为只有单个核苷酸的产物。mRNA Cleaver MicroApp支持直接添加经过修饰的核苷酸。
MAP Sequence应用程序(图3)数据处理所需的简单处理参数包括:色谱保留时间(RT)范围、最低同位素响应以及用于关联预测和实测寡核苷酸产物的质量精度容差。图4显示了MAP Sequence处理后获得的一部分结果。将实验测得的单同位素寡核苷酸产物与预测的单同位素质量数匹配后,获得了独特(绿色圆圈)和模糊(琥珀色三角形)的寡核苷酸产物分配结果。
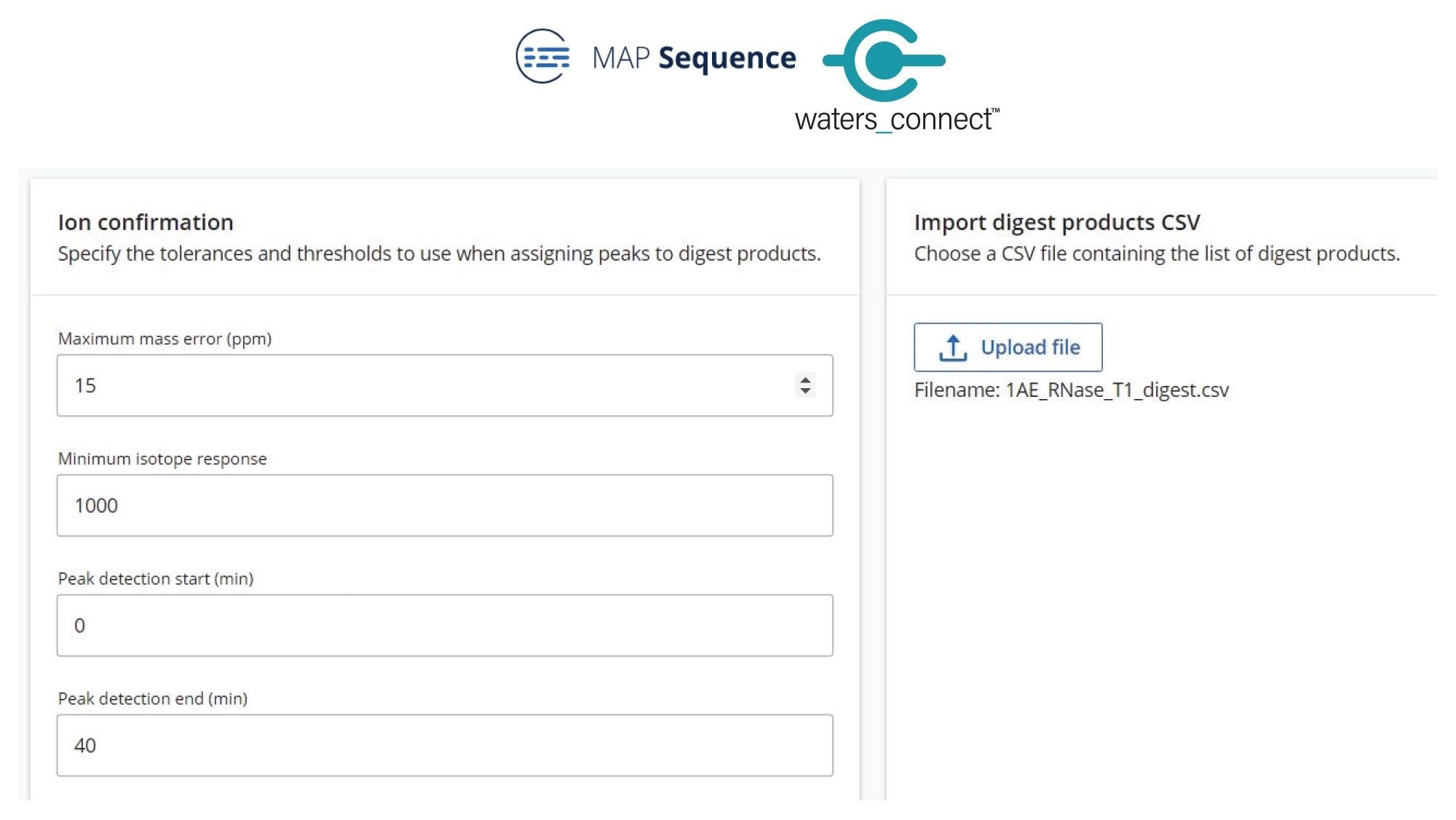 图3.MAP Sequence应用程序处理参数。此应用程序使用mRNA Cleaver软件模拟预测的寡核苷酸产物(参见右侧的csv文件)匹配酶解样品的LC-MSE产物图谱数据集。
图3.MAP Sequence应用程序处理参数。此应用程序使用mRNA Cleaver软件模拟预测的寡核苷酸产物(参见右侧的csv文件)匹配酶解样品的LC-MSE产物图谱数据集。
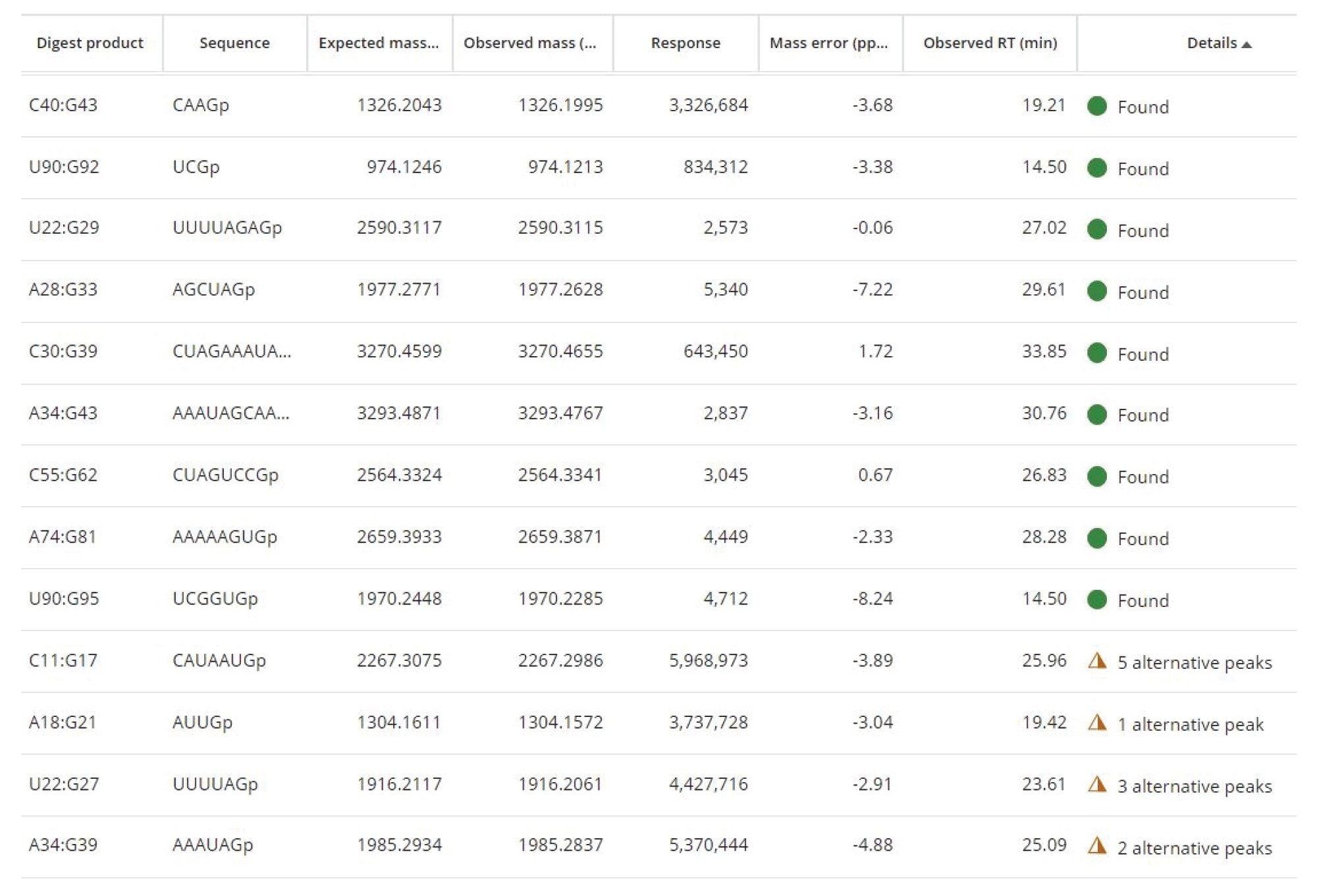 图4.MAP Sequence应用程序屏幕截图,显示了通过匹配实验测得的单同位素寡核苷酸母离子与预测的单同位素质量数获得的独特(绿色圆圈)和模糊(琥珀色三角形)寡核苷酸产物分配结果。
图4.MAP Sequence应用程序屏幕截图,显示了通过匹配实验测得的单同位素寡核苷酸母离子与预测的单同位素质量数获得的独特(绿色圆圈)和模糊(琥珀色三角形)寡核苷酸产物分配结果。
一组酶的图谱分析结果
RNase T1 sgRNA酶解物的总离子流色谱图(TIC)如图5所示。在该图中,mRNA Cleaver MicroApp预测了22种寡核苷酸产物,假设所有可能的酶切均产生含有末端鸟苷(G)残基的寡核苷酸产物。如图所示,有3对相同的寡核苷酸产物(用琥珀色标记的色谱峰突出显示,序列:AG - 峰5/18、UG - 峰15/21和CUAG - 峰6/11)产生模糊的序列分配结果。这一缺点说明了RNase T1的潜在局限性:该酶缺乏特异性,即使面对相对较短的底物(如100 mer sgRNA),也无法产生完全独特的产物。显然,如果在每个G残基之后剪切(考虑到RNA中只有4个未修饰的核苷酸,理论剪切率为25%),会生成过多的预测寡核苷酸产物,并且随着RNA序列长度增加,出现相同寡核苷酸序列产物的可能性也会显著提高。
图5汇总了MAP Sequence分配的结果:属于独特寡核苷酸产物的所有色谱峰用绿色标记,模糊序列用琥珀色标记,图片左侧显示了sgRNA序列,缺失的(不匹配的)产物用红色表示。在此色谱图中检出的大多数高丰度色谱峰都被分配为预测产物。除了上文提到的3对相同的产物外,唯一的另一个模糊产物是第一个(5'-端)寡核苷酸产物(OMEA* OMEA* OMEA* UCC UCA G)。由于前3个核苷酸的硫代磷酸化连接子中存在硫的手性中心,因此该产物可能包含多种非对映异构体。唯一缺失的(不匹配的)寡核苷酸产物是在所用的IP-RP条件下无法在色谱柱上保留的单个G残基(产物编号10、16、20)。在sgRNA的整个100 nt组成中,单核苷酸G残基是唯一未在LC-MSE数据集中检出的残基,包含至多两个RNase T1漏切并不能改善该结果。但值得注意的是,使用DPA离子对试剂能够分离两种二核苷酸物质(UG - 峰对15/21和AG - 峰对5/18),如图5所示。
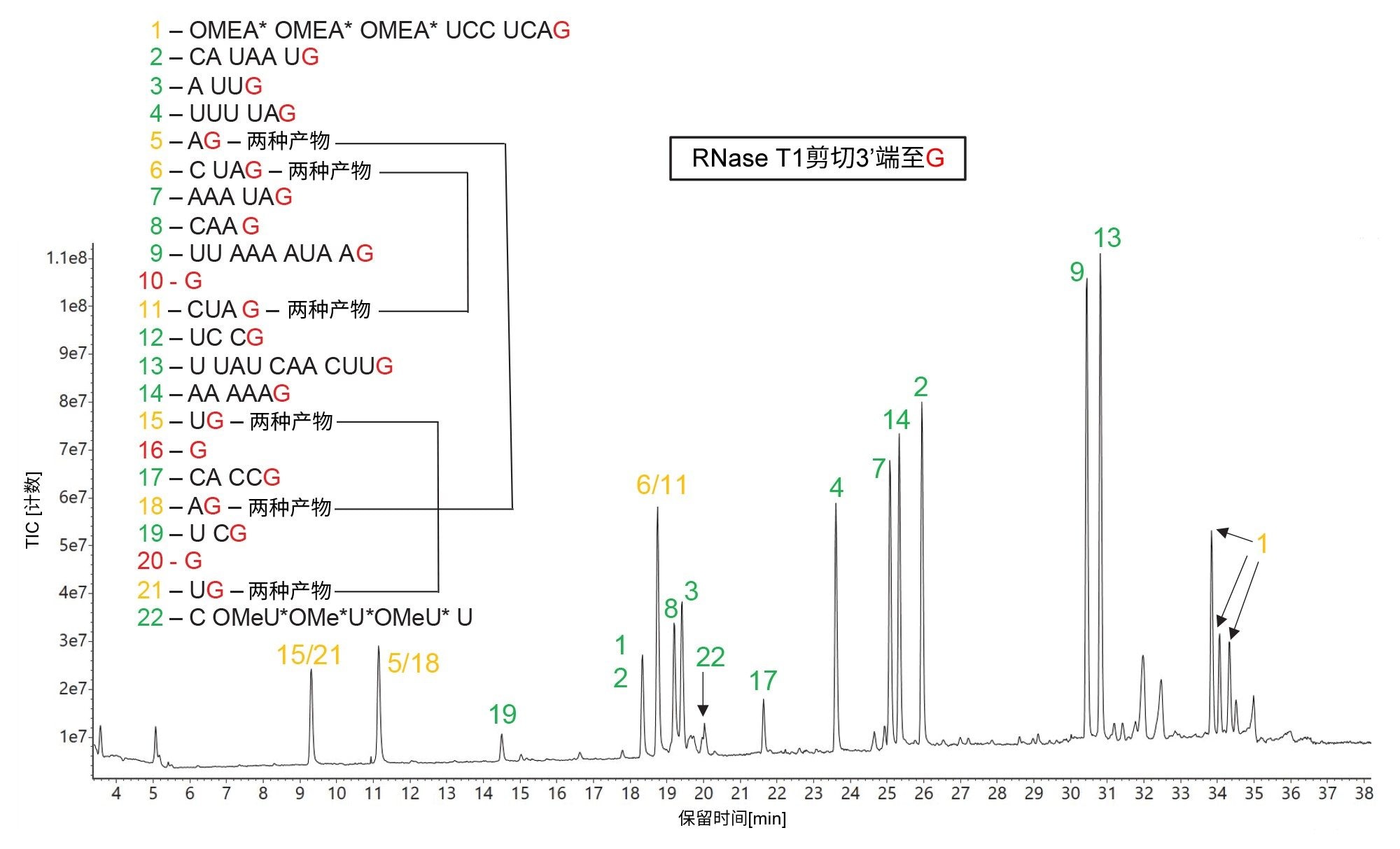 图5.RNase T1酶解HPRT1 sgRNA的TIC色谱图。预测的寡核苷酸产物根据图中左侧显示的序列进行鉴定,将用于标记检出的所有主要色谱峰。峰鉴定结果用绿色表示独特序列,用琥珀色显示的峰标签表示存在模糊(相同)序列。100 mer sgRNA的整个序列中,只有三种单核苷酸预测产物(纯G产物10、16、20)未检出。
图5.RNase T1酶解HPRT1 sgRNA的TIC色谱图。预测的寡核苷酸产物根据图中左侧显示的序列进行鉴定,将用于标记检出的所有主要色谱峰。峰鉴定结果用绿色表示独特序列,用琥珀色显示的峰标签表示存在模糊(相同)序列。100 mer sgRNA的整个序列中,只有三种单核苷酸预测产物(纯G产物10、16、20)未检出。
除RNase T1酶解外,我们还使用hRNase4(一种最近市售的酶)单独酶解了HPRT1 sgRNA7。 与RNase T1不同,hRNase4是一种尿苷(U)内切核糖核酸酶,只在U残基之后紧随嘌呤(A和G)残基的情况下剪切。在mRNA Cleaver MicroApp中定义hRNase4的两个剪切位点为U_A和U_G,这意味着剪切RNA底物的理论几率为12.5%,是RNase T1理论剪切概率的一半。图6中同一sgRNA底物hRNase4酶解物的TIC色谱图证实了这一观察结果。预测寡核苷酸产物的数量与RNase T1酶解相比显著减少(14 vs 22),与随机100 mer RNA序列剪切的预测寡核苷酸产物预期数量(12-13个产物)良好匹配。虽然不存在相同的预测序列,但与RNase T1一样,hRNase4产生了一对6 mer结构异构体(峰对5/9,AG AGC U vs A AGG CU),导致了模糊的序列分配,因为它们在所用的IP-RP条件下(与RNase T1酶解物所用的分离条件相同)未实现色谱分离。对于这种模糊分配的情况,可通过调整两种6 mer寡核苷酸产物的色谱分离条件,并使用其相应的碎片离子(高能量MSE碎片离子数据)来解决,以使用waters_connect CONFIRM Sequence应用程序分配正确序列 8
。
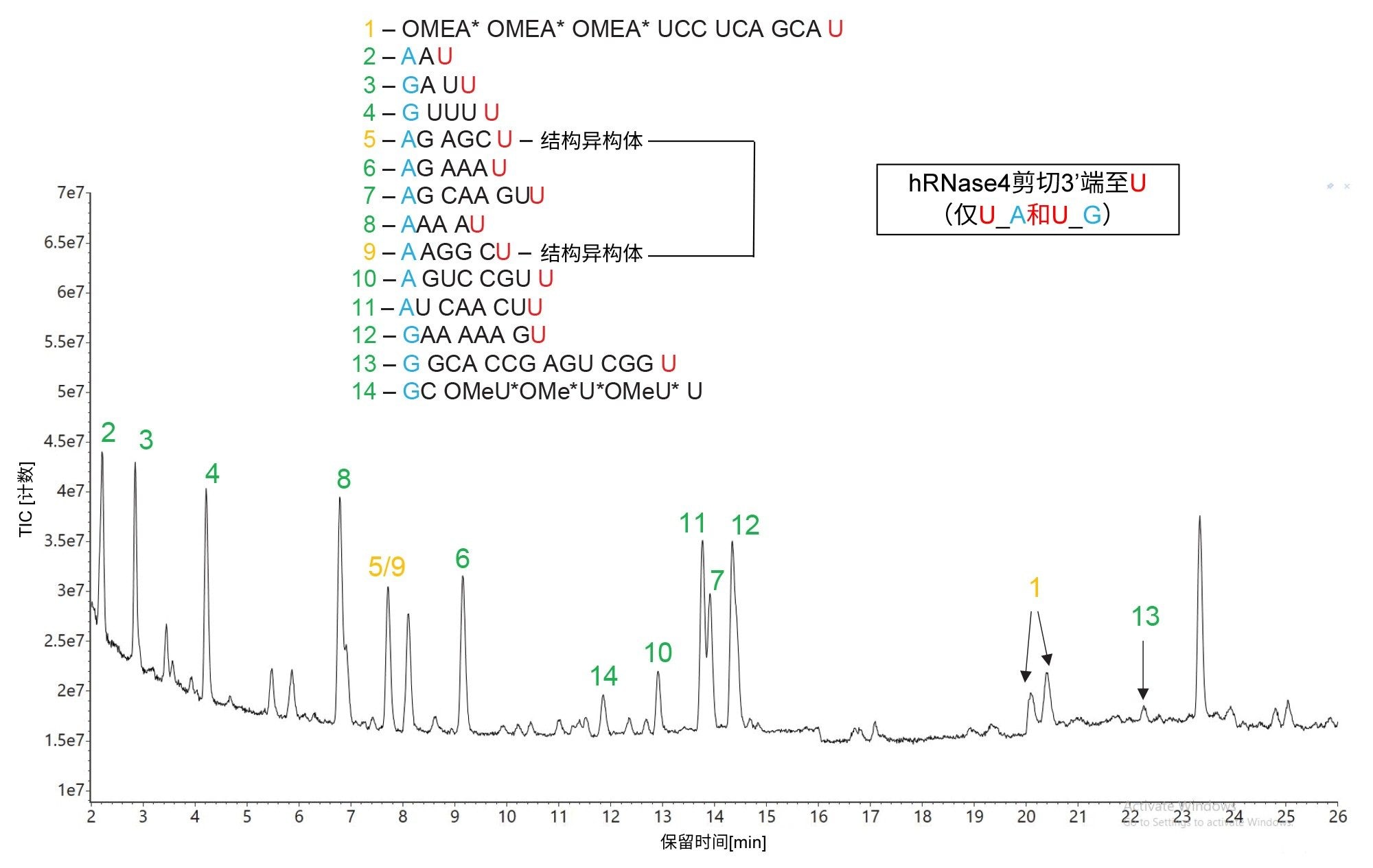 图6.hRNase 4酶解HPRT1 sgRNA的TIC色谱图。预测的寡核苷酸产物根据图中央显示的序列进行鉴定,将用于标记检出的所有主要色谱峰。峰鉴定结果用绿色表示独特序列,用琥珀色显示的峰标签表示存在模糊(相同)序列。只有一个预测寡核苷酸产物(峰13)未检出。
图6.hRNase 4酶解HPRT1 sgRNA的TIC色谱图。预测的寡核苷酸产物根据图中央显示的序列进行鉴定,将用于标记检出的所有主要色谱峰。峰鉴定结果用绿色表示独特序列,用琥珀色显示的峰标签表示存在模糊(相同)序列。只有一个预测寡核苷酸产物(峰13)未检出。
唯一的另一个模糊分配是由一个晚洗脱的双峰引起的,该双峰对应于第一个5'-端寡核苷酸产物(OMEA* OMEA* OMEA* UCC UCA GCA U),该产物由于具有多个手性中心也表现出异质性。低丰度寡核苷酸产物(相对于最高丰度产物的ESI-MS谱图强度,丰度为0.6%)的ESI-MS谱图如图7所示。在该谱图中检测到的双电荷和三电荷寡核苷酸母离子证实存在14 mer预测寡核苷酸产物(序列:G GCA CCG AGU CGG U),在MAP Sequence中将离子同位素响应阈值降低至1000计数后检出该产物。关于RNase T1和hRNase4的生物活性,最终总结如下:两种酶均在其偏好的残基(G/U残基的3'-侧)后剪切,而RNase T1在酶解寡核苷酸产物的3'-端添加了线性磷酸酯,hRNase4产生的酶解产物则包含了水平大致相等的3'-端线性磷酸酯和环状磷酸酯,这一点通过软件检测得出。RNase T1和hRNase4都能完全剪切RNA底物,目前的TIC色谱图中没有证据表明存在未酶解的sgRNA(数据未显示)。
RapiZyme MC1和RapiZyme Cusativin是沃特世公司最近推出的两种新酶。MC1具有较高的尿苷特异性,而Cusativin具有较高的胞苷特异性9,10。
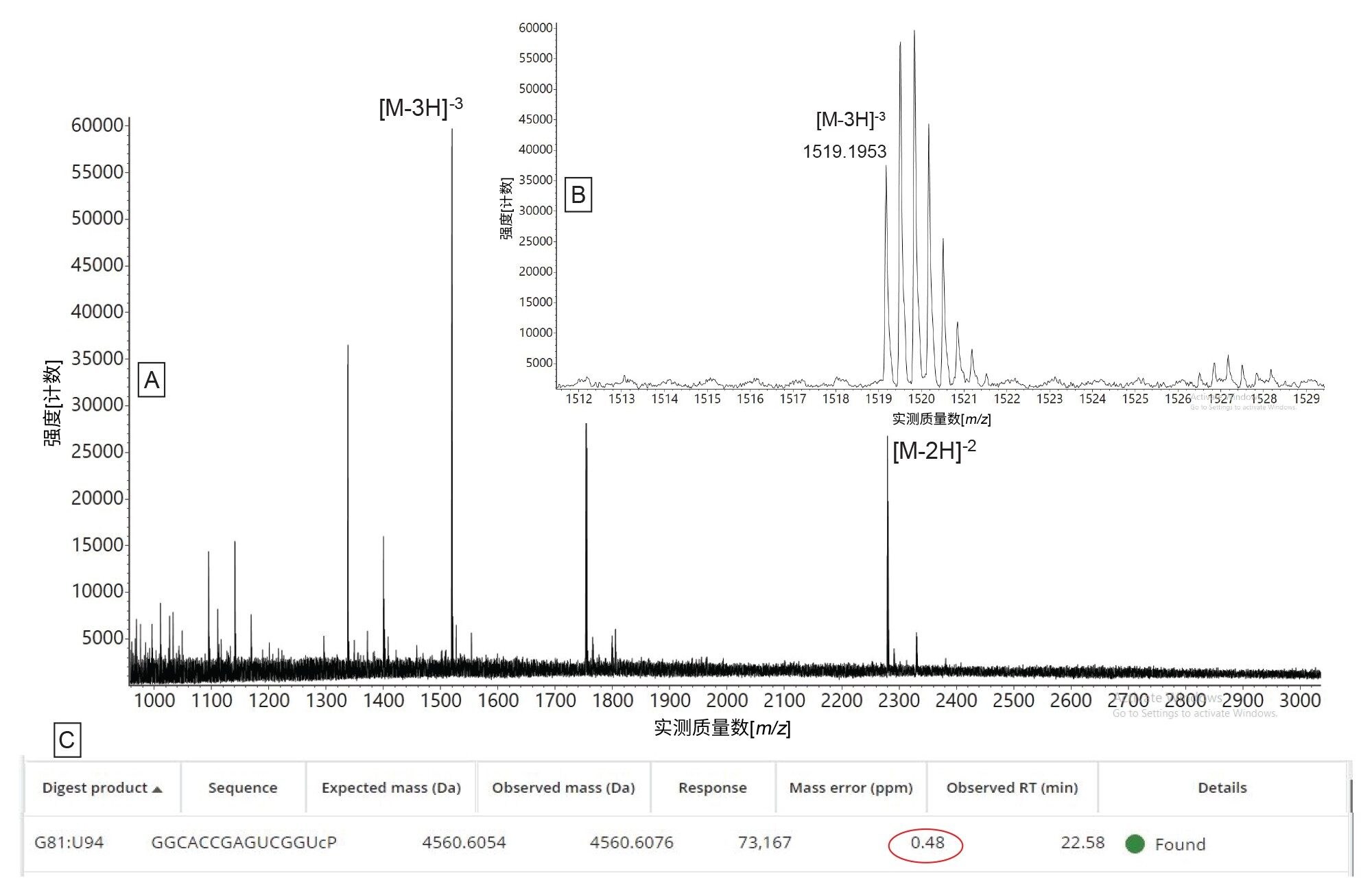 图7.MAP Sequence应用程序自动分配使用hRNase4酶酶解HPRT1 sgRNA后生成的低强度寡核苷酸产物:(A)寡核苷酸产物G GCA CCG AGU CGG U的ESI-MS合并谱图;(B)插图显示了三电荷寡核苷酸母离子的同位素分布;(C) MAP Sequence应用程序结果,显示了将ESI谱图中的双电荷和三电荷母离子分配给该寡核苷酸产物,质量数误差小于1 ppm。
图7.MAP Sequence应用程序自动分配使用hRNase4酶酶解HPRT1 sgRNA后生成的低强度寡核苷酸产物:(A)寡核苷酸产物G GCA CCG AGU CGG U的ESI-MS合并谱图;(B)插图显示了三电荷寡核苷酸母离子的同位素分布;(C) MAP Sequence应用程序结果,显示了将ESI谱图中的双电荷和三电荷母离子分配给该寡核苷酸产物,质量数误差小于1 ppm。
图8显示了HPRT1 sgRNA底物的MC1酶解物的LC-MS分析TIC色谱图,以及MAP Sequence处理后得到的相应序列分配结果。由于MC1有五个不同的预期酶切位点(A_U / C_U / U_U / C_A / C_G),同时还考虑了至多两个漏切,因此mRNA Cleaver应用程序会产生数量相对较多的预测寡核苷酸产物。实验数据中未检出许多预测的短链寡核苷酸产物(其中包含若干单核苷酸和二核苷酸),而大多数序列覆盖率分配所用的寡核苷酸产物都包含漏切位点。这类似于胰蛋白酶(高特异性)和胃蛋白酶(低特异性)对蛋白质的酶解。前者可生成较简单的肽图,后者可能生成序列覆盖范围重叠的图。
另外值得一提的是,与RNase T1和hRNase4不同,MC1在其特定靶标之前(U残基的5'-端)剪切,并且主要将一个环状磷酸酯添加到寡核苷酸酶解产物的3’-端。Cusativin主要在胞苷之后(3'-端至C残基)剪切,但它还有其他四个次要的酶切位点。总体而言,Cusativin预期有七个不同的酶切位点,在mRNA Cleaver MicroApp中定义为:C_U / C_A / C_G / U_U / A_U / U_A / G_U。与MC1一样,Cusativin主要在每个寡核苷酸酶解产物的3'-端添加一个环状磷酸酯,尽管在软件中检测到两种形式(环状和线性)。
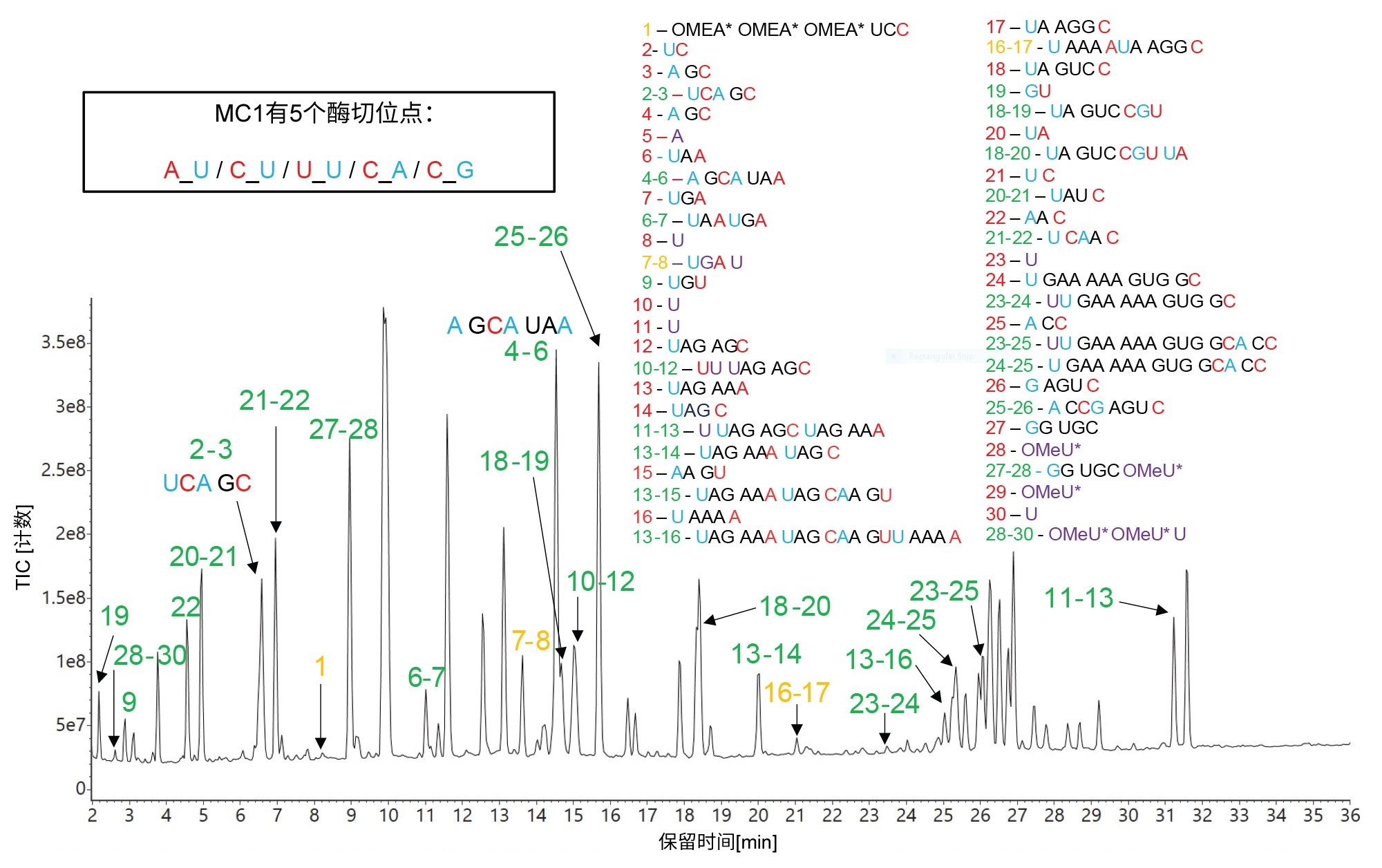 图8.RapiZyme MC1酶解HPRT1 sgRNA的TIC色谱图。预测的寡核苷酸产物根据图中右侧两张图中的序列进行鉴定,将用于标记检出的所有色谱峰。峰鉴定结果用绿色表示独特序列,用琥珀色显示的峰标签表示存在模糊(相同)序列。大部分序列分配是由具有一两个漏切位点的寡核苷酸产物进行的。
图8.RapiZyme MC1酶解HPRT1 sgRNA的TIC色谱图。预测的寡核苷酸产物根据图中右侧两张图中的序列进行鉴定,将用于标记检出的所有色谱峰。峰鉴定结果用绿色表示独特序列,用琥珀色显示的峰标签表示存在模糊(相同)序列。大部分序列分配是由具有一两个漏切位点的寡核苷酸产物进行的。
Cusativin的特异性在HPRT1 sgRNA酶解物的TIC色谱图中有所体现(图9),因为许多分配的寡核苷酸产物都含有漏切位点。为了覆盖sgRNA序列中可能无法检出的部分,具有相对较多的漏切位点显然是有利的,因为不含漏切位点的相应产物长度都很短(单核苷酸、二核苷酸或三核苷酸),并且难以通过IP-RP UPLC方法分离。MC1和Cusativin都能完全剪切RNA底物,目前未酶解sgRNA的TIC色谱图中没有证据表明这一点(数据未显示)。RapiZyme MC1和RapiZyme Cusativin在另一篇应用纪要中重点介绍,对这些独特的酶及其在RNA酶解方面的应用提供了更多详细信息11。
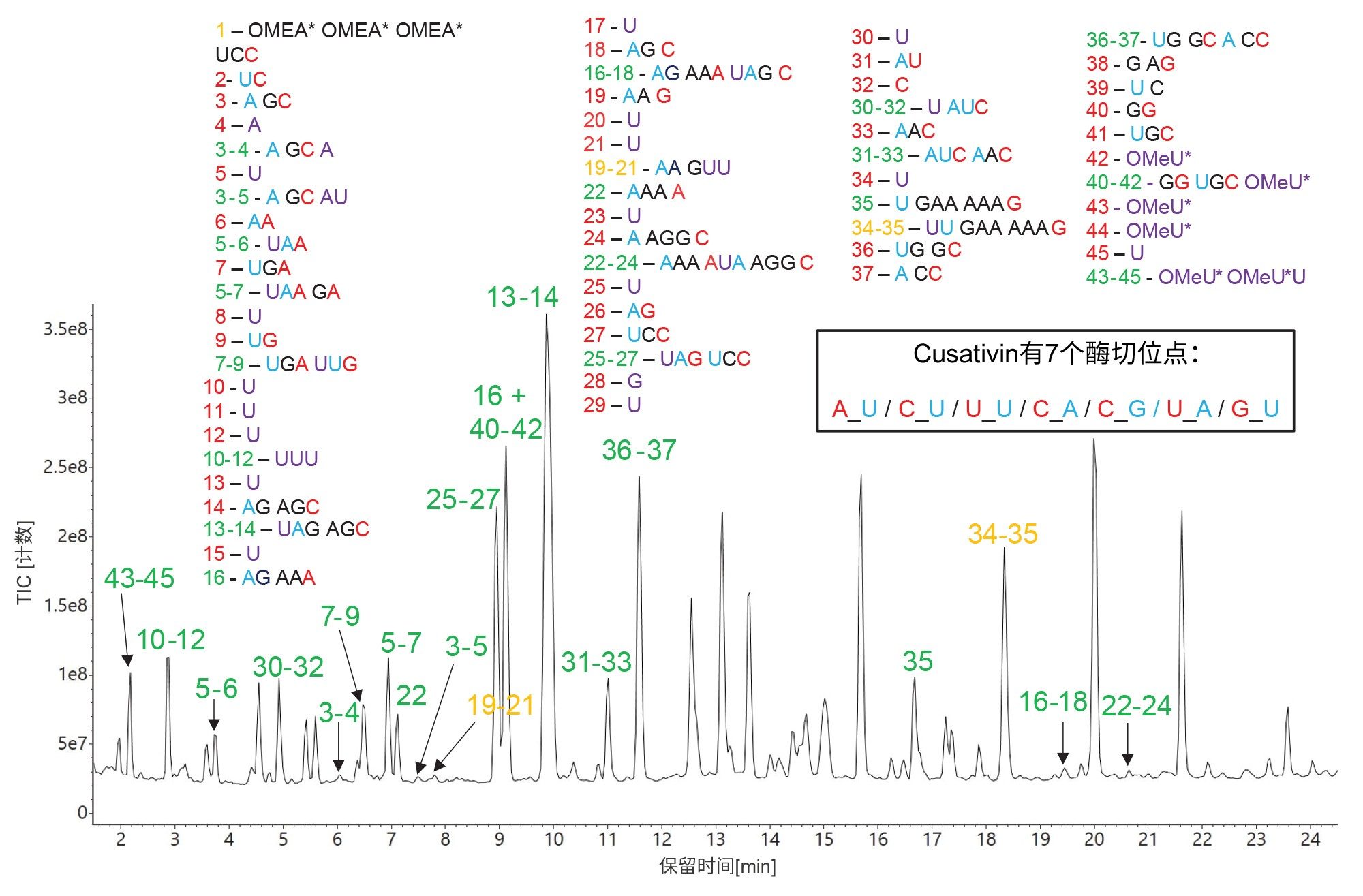 图9.RapiZyme Cusativin酶解HPRT1 sgRNA的TIC色谱图。预测的寡核苷酸产物根据图中顶部四张图中的序列进行鉴定,将用于标记检出的所有色谱峰。峰鉴定结果用绿色表示独特序列,用琥珀色显示的峰标签表示存在模糊(相同)序列。大部分序列分配是由具有一两个漏切位点的寡核苷酸产物进行的。
图9.RapiZyme Cusativin酶解HPRT1 sgRNA的TIC色谱图。预测的寡核苷酸产物根据图中顶部四张图中的序列进行鉴定,将用于标记检出的所有色谱峰。峰鉴定结果用绿色表示独特序列,用琥珀色显示的峰标签表示存在模糊(相同)序列。大部分序列分配是由具有一两个漏切位点的寡核苷酸产物进行的。
基于上述观察结果,增加MC1和Cusativin的漏切数量,允许至多3个漏切,尝试使这些酶获得完全的序列覆盖率。此外,还扩展了mRNA Cleaver中定义的酶切位点,包括sgRNA序列中存在的几个修饰残基周围的酶切位点。例如,对于MC1,增加了三个酶切位点:C_Y / Y_Y / Y_U,其中Y表示2'-OMe硫代磷酸化尿苷。这样做的结果是,两种酶解物在使用MAP Sequence处理后,都检测到一种新预测的寡核苷酸产物,序列为YYU。使用CONFIRM Sequence应用程序处理高能量碎片离子谱图后,确认了该产物的鉴定结果8。 其单电荷母离子的同位素分布以及CONFIRM Sequence应用程序生成的点图如图10所示。该三核苷酸的序列经过验证,表明MC1和Cusativin均在经过修饰的尿苷残基之间剪切(Y_Y基序剪切),这是一种独特且有用的酶功能。hRNase4酶无法剪切经修饰的尿苷,但MC1和Cusativin不受2'-端尿苷修饰影响,因此能够产生独特的酶解产物7。
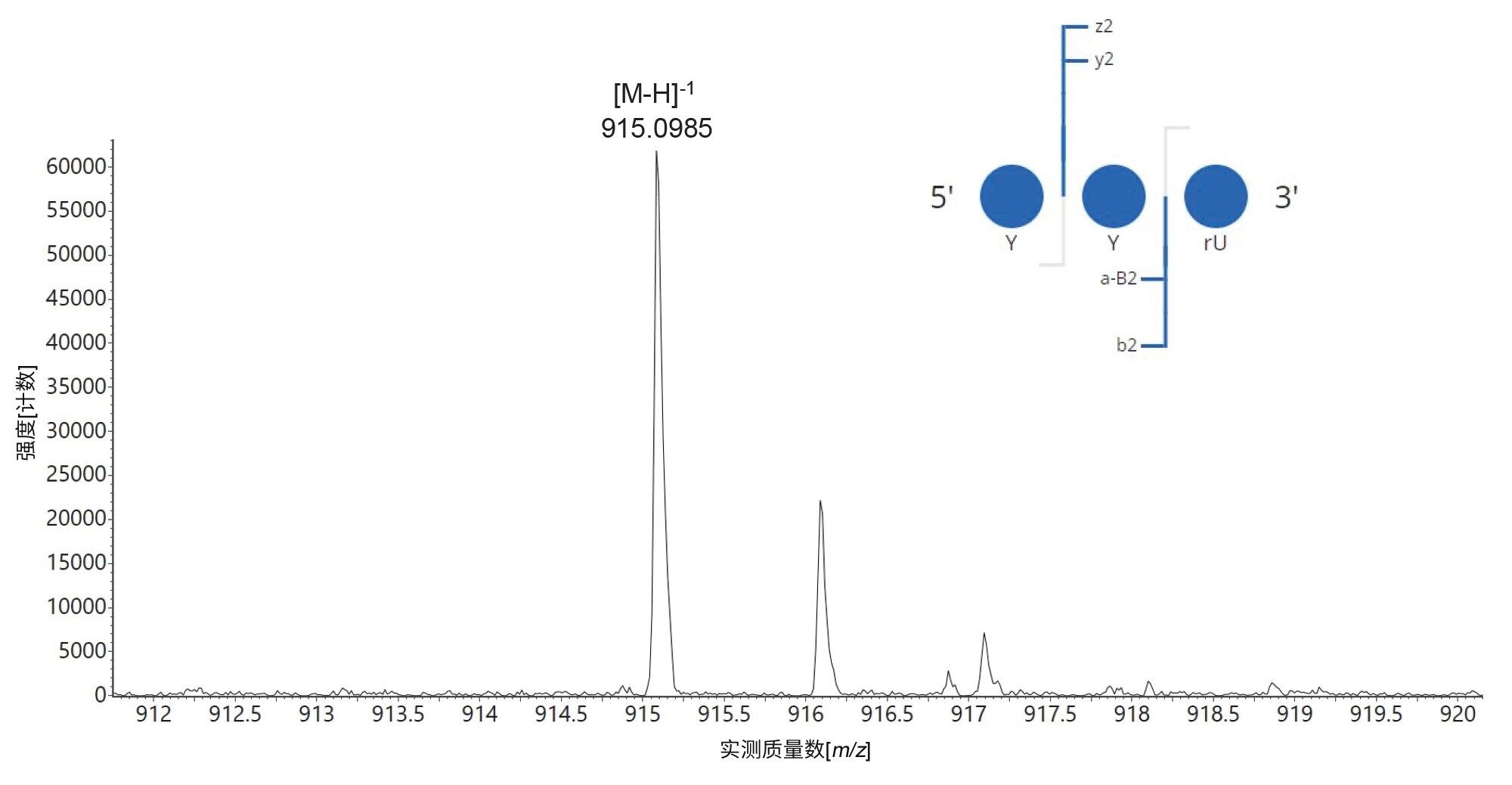 图10.在经修饰sgRNA的RapiZyme MC1酶解物中检测到的三核苷酸YYU单电荷离子的同位素分布。插图为CONFIRM Sequence应用程序的界面截图,显示了碎片离子点图,确认该寡核苷酸酶解产物的序列为YYU(100 mer sgRNA序列中的98-100号残基)。这些结果证实,MC1具有切割2'-端修饰尿苷周围的独特能力,这是其他核糖核酸酶所缺少的功能。
图10.在经修饰sgRNA的RapiZyme MC1酶解物中检测到的三核苷酸YYU单电荷离子的同位素分布。插图为CONFIRM Sequence应用程序的界面截图,显示了碎片离子点图,确认该寡核苷酸酶解产物的序列为YYU(100 mer sgRNA序列中的98-100号残基)。这些结果证实,MC1具有切割2'-端修饰尿苷周围的独特能力,这是其他核糖核酸酶所缺少的功能。
在RNA分析工作流程的最后一步(图1),使用Coverage Viewer MicroApp(图11)生成RNase T1、hRNase4、RapiZyme MC1和RapiZyme Cusativin酶解相同sgRNA获得的序列覆盖率汇总。RNase T1和hRNase4的序列覆盖率分别在80~90%范围内,而MC1和Cusativin酶解对该分子的覆盖率达到了100%。总体而言,这些结果表明,当结合多种酶的结果时,可以在寡核苷酸产物图谱分析中获得更高的可信度。
 图11.Coverage Viewer MicroApp结果,汇总了RNase T1、hRNAase4、RapiZyme MC1和RapiZyme Cusativin酶解后各自获得的序列覆盖率。
图11.Coverage Viewer MicroApp结果,汇总了RNase T1、hRNAase4、RapiZyme MC1和RapiZyme Cusativin酶解后各自获得的序列覆盖率。
利用质谱技术对大分子RNA(sgRNA、mRNA)进行序列确认,仍然是一个正在积极开发的研究领域。为处理这些复杂数据集开发自动化软件,以及本文探讨的更庞大的酶工具集,是提高这种新兴方法实用性的关键。
结论
- 本研究展示了新的软件工具,可通过使用多种酶酶解sgRNA得到的UPLC-MS数据,高效绘制RNA酶解产物的图谱。利用新的RNA酶解产物图谱分析应用程序(MAP Sequence)成功处理了每种酶的sgRNA酶解物数据,生成产物图谱,用于确认预测序列。
- 两种新型的RNase T2酶(RapiZyme MC1和RapiZyme Cusativin)产生了更多的寡核苷酸酶解产物和漏切位点,表现出独特的酶解特异性。这些酶在获取具有重叠序列区域的寡核苷酸产物图谱方面具有潜在作用。
- 所有酶均获得了较高的序列覆盖率,MC1和Cusativin在该分子的寡核苷酸酶解产物水平上实现了100%序列确认。实际上,并非所有sgRNA分子以及更长的mRNA序列都能达到这样的效果。将一组酶解结果相结合,可提高对质量指纹图谱分析方法准确性的整体信心,有机会为RNA类治疗药物实现完全的序列覆盖率。
- 虽然寡核苷酸图谱分析目前主要用于开发阶段,但合规的waters_connect信息学平台上一体化的UPLC-MS数据采集和数据处理工作流程在生产和质控方面也有应用潜力。
参考资料
- Jinek M, Chylinsky K, Fonfara I, Hauer M, Doudna JA, Charpentier E. A Programmable Dual-RNA-Guided DNA Endonuclease in Adaptive Bacterial Immunity, Science, 2012, 337, 816–821.
- Jiang F, Doudna JA.CRISPR-Cas9 Structures and Mechanisms, Annu Rev Biophys, 2017, 46, 505–529.
- Ganbaatar U, Liu C. CRISPR-Based COVID-19 Testing: Toward Next Generation Point of Care Diagnostics, Front Cell Infect Microbiol, 2021, 11.https://doi.org/10.3389/fcimb.2021.663949
- Catalin E. Doneanu, Patrick Boyce, Henry Shion, Joseph Fredette, Scott J. Berger, Heidi Gastall, Ying Qing Yu.使用搭载ACQUITY Premier的BioAccord系统和全新的自动化INTACT Mass应用程序对siRNA、单向导RNA和杂质进行LC-MS分析.沃特世应用纪要.720007546ZH.2022年
- Goyon A, Scott B, Kurita K, Crittenden CM, Shaw D, Lin A, Yehl P, Zhang K. Full Sequencing of CRISPR/Cas9 Single Guide RNA (sgRNA) via Parallel Ribonuclease Digestions and Hydrophilic Interaction Liquid Chromatography High-Resolution Mass Spectrometry Analysis, Anal Chem, 2022, 93, 14792–14801. doi: 10.1021/acs.analchem.1c03533
- Rebecca J. D'Esposito, Catalin E, Doneanu Heidi Gastall, Scott J. Berger, Ying Qing Yu.使用BioAccord LC-MS系统和INTACT Mass waters_connect应用程序进行RNA CQA分析.沃特世应用纪要.720008130ZH.2023年.
- Wolf EJ, Grunberg S, Dai N, Chen T-H, Roy B, Yigit E, Correa IR.Human RNase 4 Improves mRNA Sequence Characterization by LC-MS/MS, Nucleic Acid Res, 2022, 50, e106. DOI:10.1093/nar/gkac632
- Catalin E. Doneanu, Chris Knowles, Matt Gorton, Henry Shion, Joseph Fredette, Ying Qing Yu.CONFIRM Sequence:⼀款⽤于合成寡核苷酸及其杂质测序的waters_connect应⽤程序.沃特世应用纪要.720007677ZH.2022年.
- Grunberg S, Wolf EJ, Jin J, Ganatra MD, Becker K, Ruse C, Taron CH, Correa IR, Yigit E. Enhanced Expression and Purification of Nucleotide-specific Ribonucleases MC1 and Cusativin, Protein Expr Purif Acid Res, 2022, 190, 105987.doi:10.1016/j.pep.2021.105987
- Thakur P, Atway J, Limbach PA, Addepalli B. RNA Cleavage Properties of Nucleobase-Specific RNase MC1 and Cusativin Are Determined by the Dinucleotide-Binding Interactions in the Enzyme-Active Site, Int J Mol Sci, 2022, 23, 7021.
- Balasubrahmanyam Addepalli Tatiana Johnston, Christian Reidy, Matthew A. Lauber.Tunable Digestions of RNA Using RapiZyme™ RNases to Confirm Sequence and Map Modifications.沃特世应用纪要.720008539.2024.
720008553ZH,2024年9月