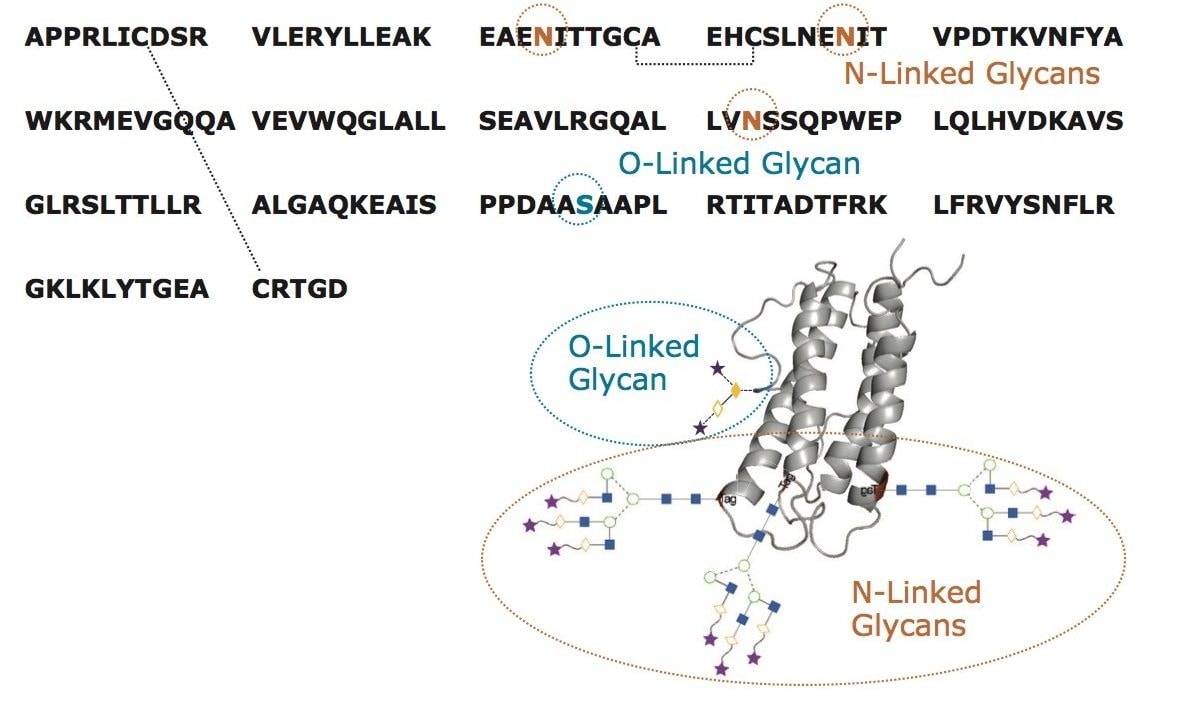
RapiFluor-MS 標識および HILIC プロファイリングを用いた rhEPO の遊離 N 結合型糖鎖分析
遺伝子組み換えヒトエポエチン(rhEPO)のグリコシル化は、過去に多数研究されています4-5, 8-13。 これまでの研究の多くは、比較的複雑な技術を必要としていました。本研究では、EPO 分析に 2 つの容易かつ相補的な LC ベースのアプローチを確立することを目的としています。1 つは N グリコシル化についての情報を、もう 1 つは O グリコシル化についての情報を提供することができるものです。
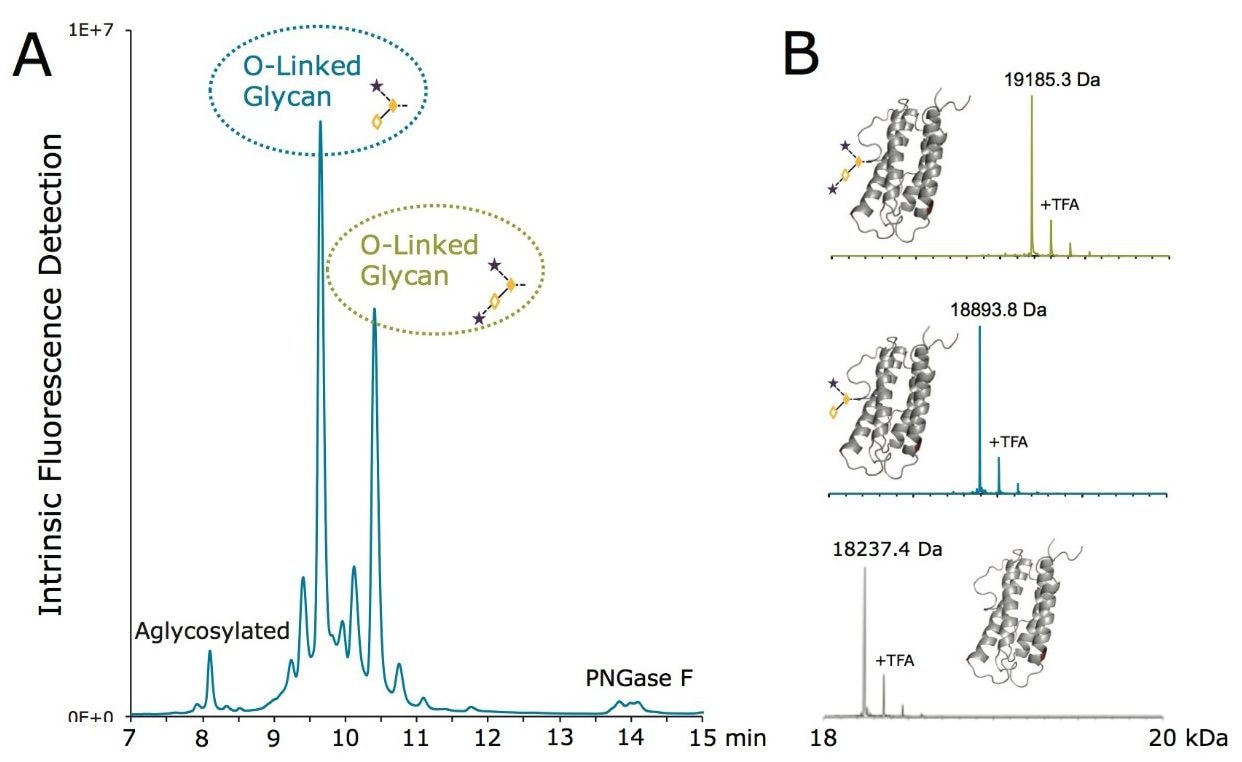
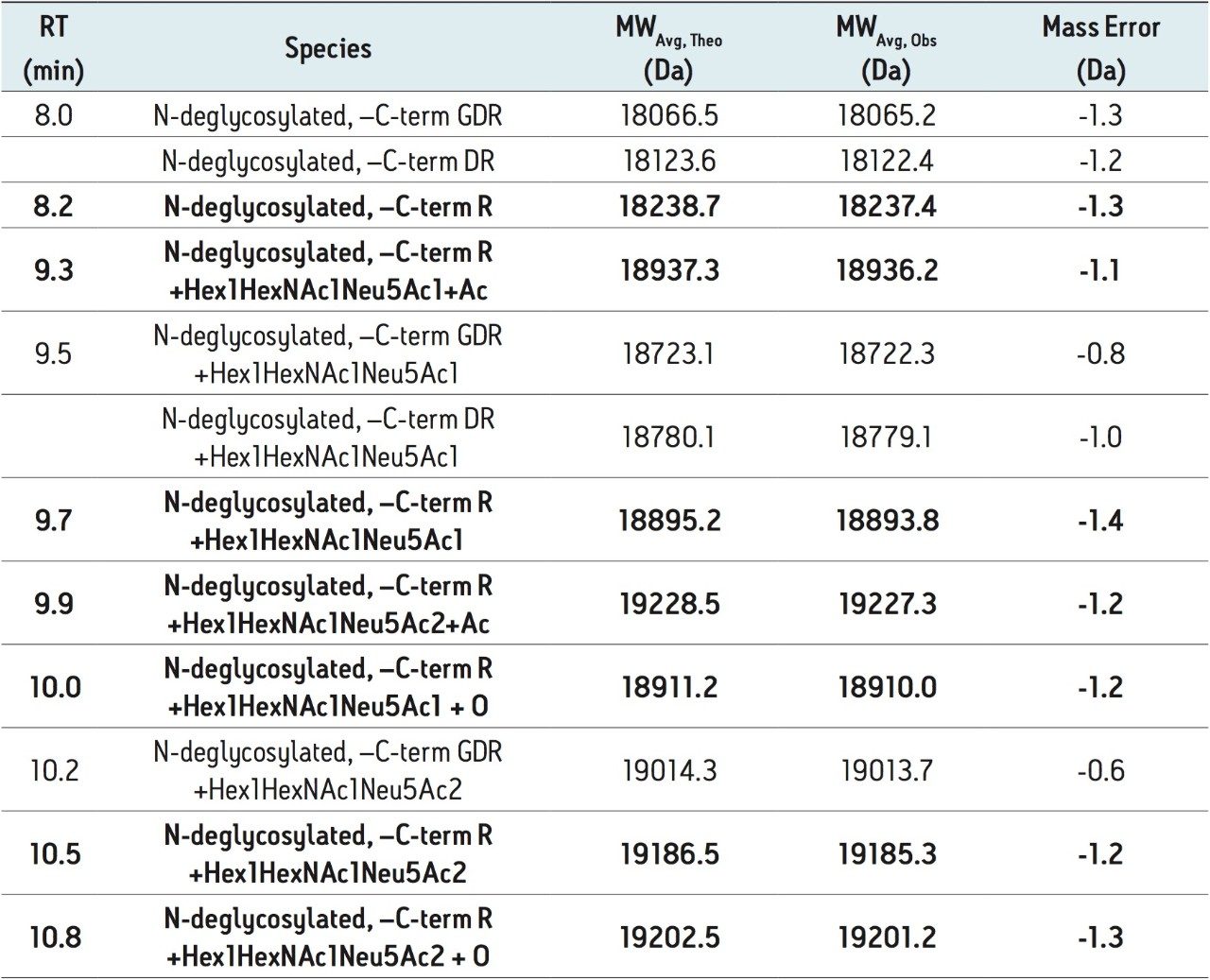
rhEPO の N 結合型糖鎖のプロファイルは、新規糖鎖標識試薬 RapiFluor-MS を含む新しいサンプル前処理ストラテジーにより迅速に得ることができます。GlycoWorks RapiFluor-MS N-Glycan キットに基づくこのサンプル前処理により、N 結合型糖鎖を迅速に切り出し、蛍光およびエレクトロスプレーイオン化質量分析(ESI-MS)検出の感度を向上するタグを標識します 14。既存のアプリケーションでは、RapiFluor-MS は主に様々な IgG サンプルの分析に用いられていました14-16。 加えて、GlycoWorks RapiFluor-MS N-Glycan キットのプロトコールを用いて、rhEPO のような高度にグリコシル化されたタンパク質からも十分にサンプルを調製することができます。
RapiFluor-MS 標識 N 結合型糖鎖は、親水性相互作用クロマトグラフィー(HILIC)に適していることが証明されています。従って、RapiFluor-MS の HILIC-蛍光-MS は、タンパク質の N グリコシル化の詳細を確認する非常に強力なツールとして登場しました14。
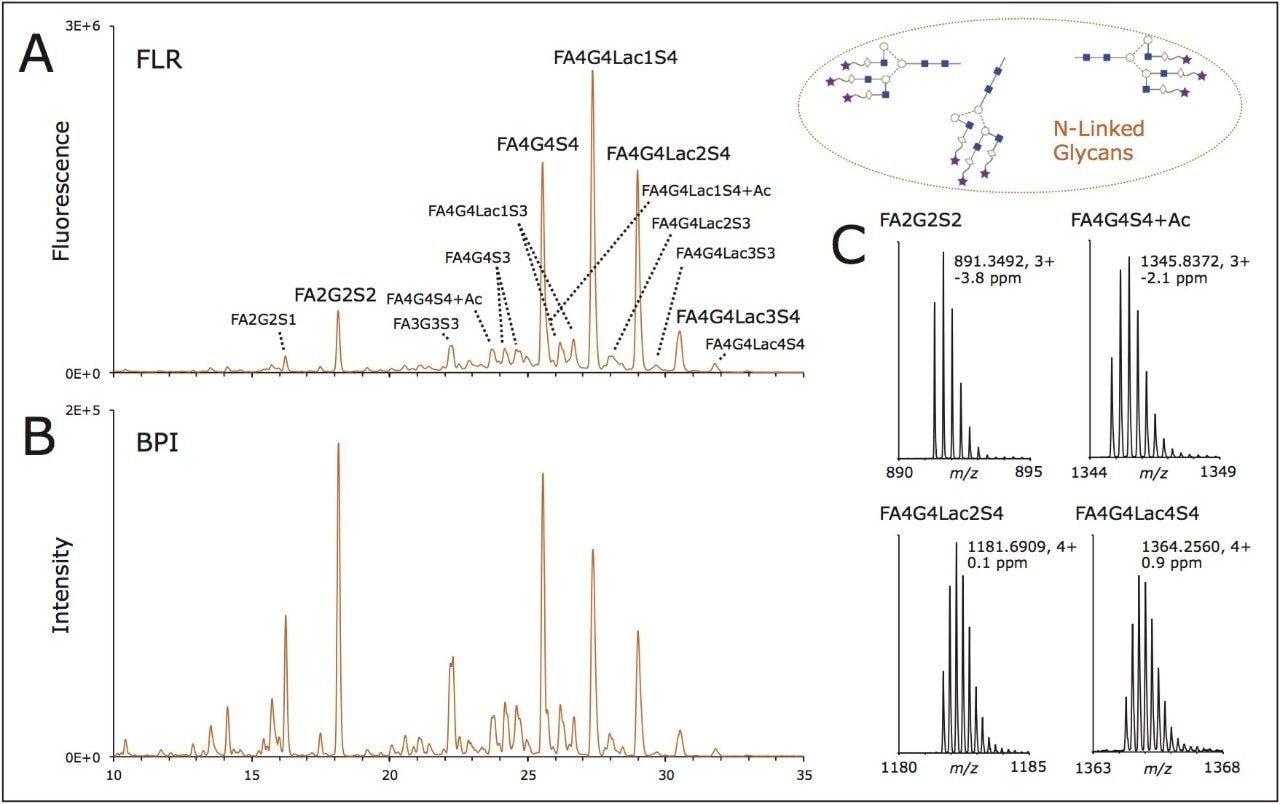
この目的のために、rhEPO から得られた RapiFluor-MS N 結合型糖鎖のサンプルは HILIC を用いてプロファイリングしました。N 結合型糖鎖の高分離を得るために、最近発表したワイドポアアミドカラム、ACQUITY UPLC Glycoprotein BEH Amide、300Å、1.7 µm カラムを本研究には選択しました。このカラムは、糖ペプチドや糖タンパク質などの高分子の HILIC 分離を促進するために設計されました。また一方で、ワイドポアパーティクルの構造は、高度に分岐した三分岐もしくは四分岐 N 結合型糖鎖分離におけるピークキャパシティを 10~20% 向上することも示されており、17 一般的に高度な分岐性を示す EPO N-結合型糖鎖の HILIC プロファイリングに最適な選択となります。図 2A には、0.4 µg の rhEPO から得られた
RapiFluor-MS N 結合型糖鎖の HILIC 蛍光および base peak intensity (BPI) MS クロマトグラムを示しました。この比較的限られた量のサンプルから、高い S/N のクロマトグラムが得られました。蛍光分析の感度により、全プロファイルに渡って正確な相対定量が可能になります。MS クロマトグラムの S/N も特筆すべき点で、N 結合型糖鎖の構造が大きくなるにつれて MS 感度は低下することにも注目してください。それにも関わらず、RapiFluor-MS 試薬をイオン輸送効率と感度が向上した新世代の MS である Xevo G2-XS QTof と組み合わせることで、このデータの品質が可能となりました。この QTof テクノロジーにより、様々な N 結合型糖鎖種の帰属に使用した図 2B に示す一連の MS スペクトルに見られるような、かつてない感度と高い MS 分解能が実現します。