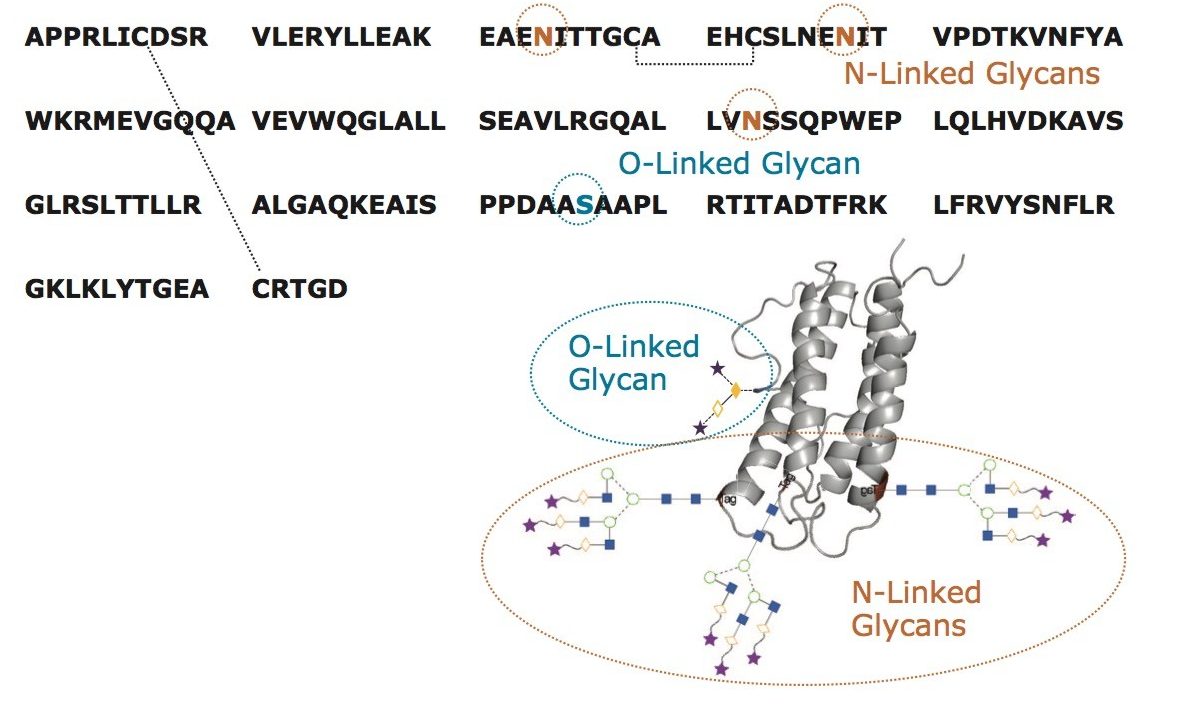
Released N-Glycan analysis of rhEPO using RapiFluor-MS labeling and HILIC profiling
The glycosylation of recombinant, human epoetin (rhEPO) has been investigated many times before.4-5, 8-13 In large part, these previous studies have required relatively involved techniques. With this work, it was our objective to establish two facile and complementary, LC based approaches for the analysis of EPO, one capable of providing information about N-glycosylation and the other information about O-glycosylation.
A profile of the N-glycans from rhEPO can be readily obtained with a new sample preparation strategy involving the novel glycan labeling reagent, RapiFluor-MS. This sample preparation, based on the GlycoWorks RapiFluor-MS N-Glycan Kit, allows an analyst to rapidly release N-glycans and label them with a tag that provides enhanced sensitivity for fluorescence and electrospray ionization mass spectrometric (ESI-MS) detection.14 In previous applications, RapiFluor-MS has been predominately used in the analysis of different IgG samples.14-16 Nevertheless, using the protocol from the GlycoWorks RapiFluor-MS N-Glycan Kit, an analyst can successfully prepare samples from even heavily glycosylated proteins, such as rhEPO.
RapiFluor-MS labeled N-glycans have proven to be amenable to hydrophilic interaction chromatography (HILIC). Accordingly, HILIC-fluorescence-MS of RapiFluor-MS has emerged as a very powerful tool for detailing the N-glycosylation of proteins.14
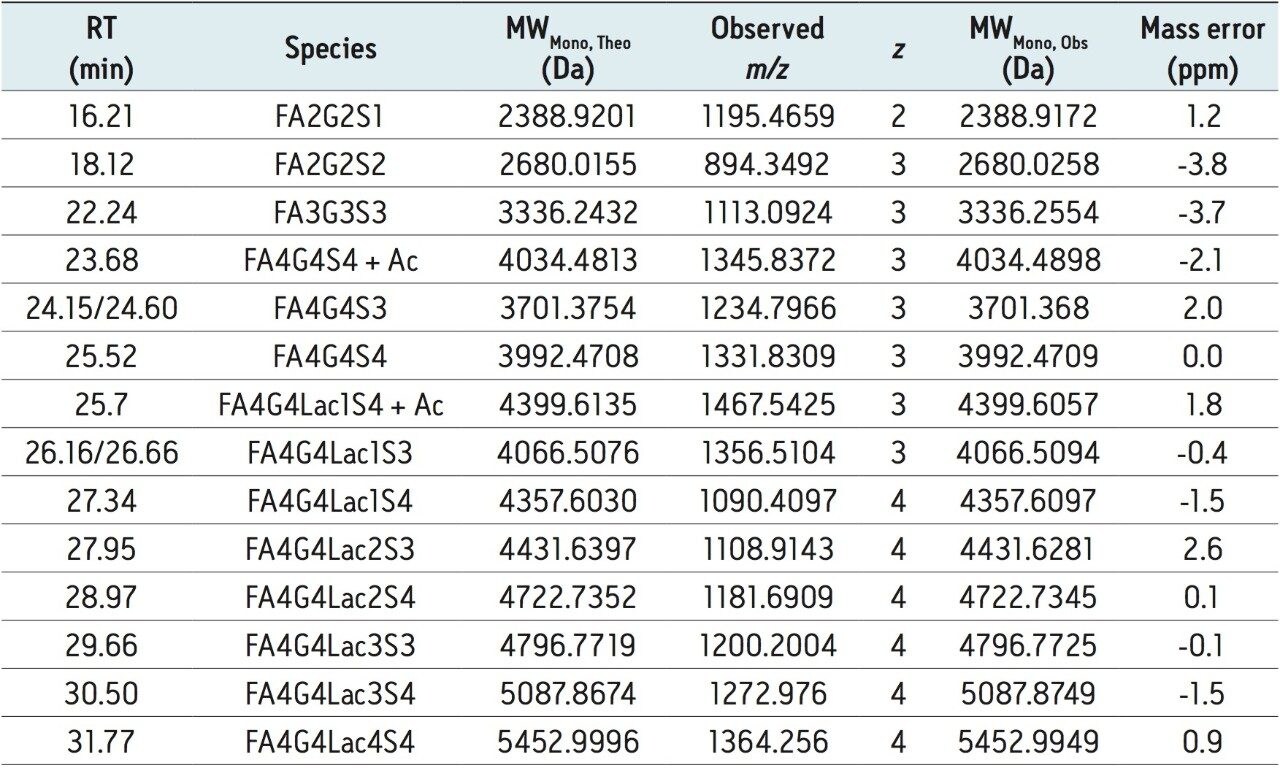
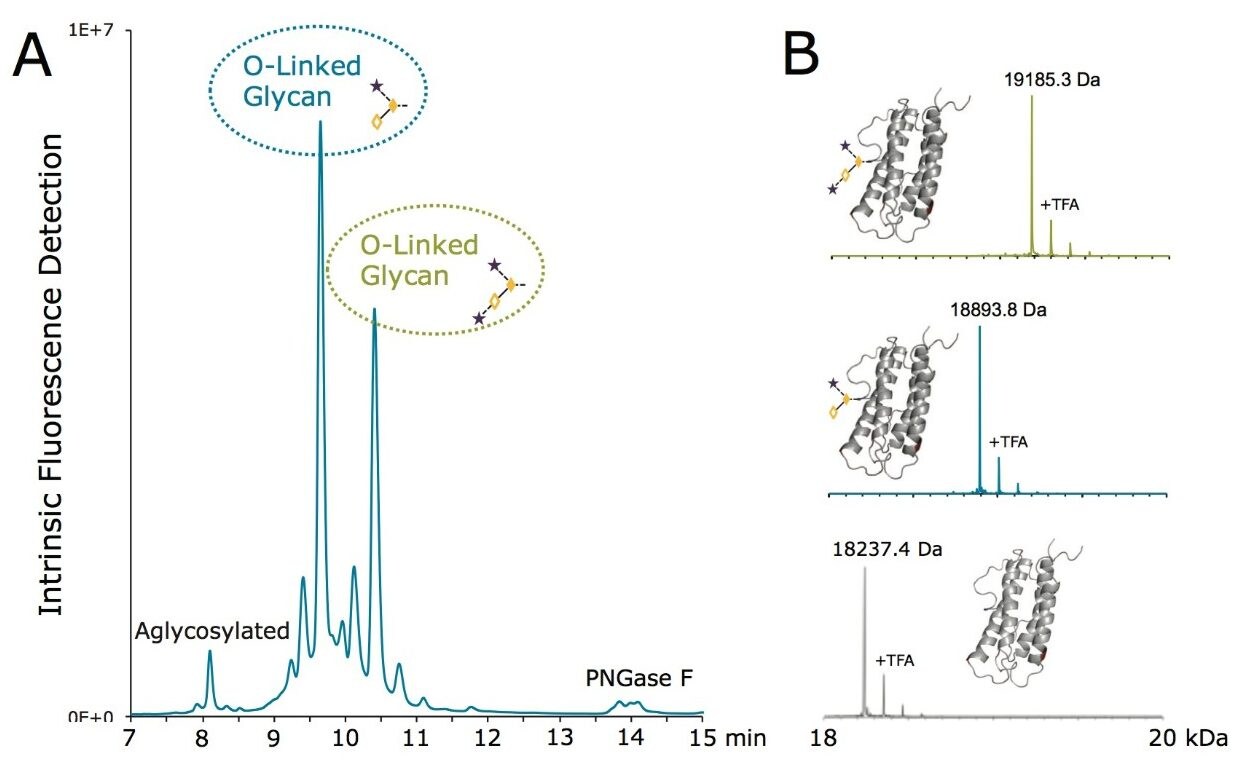
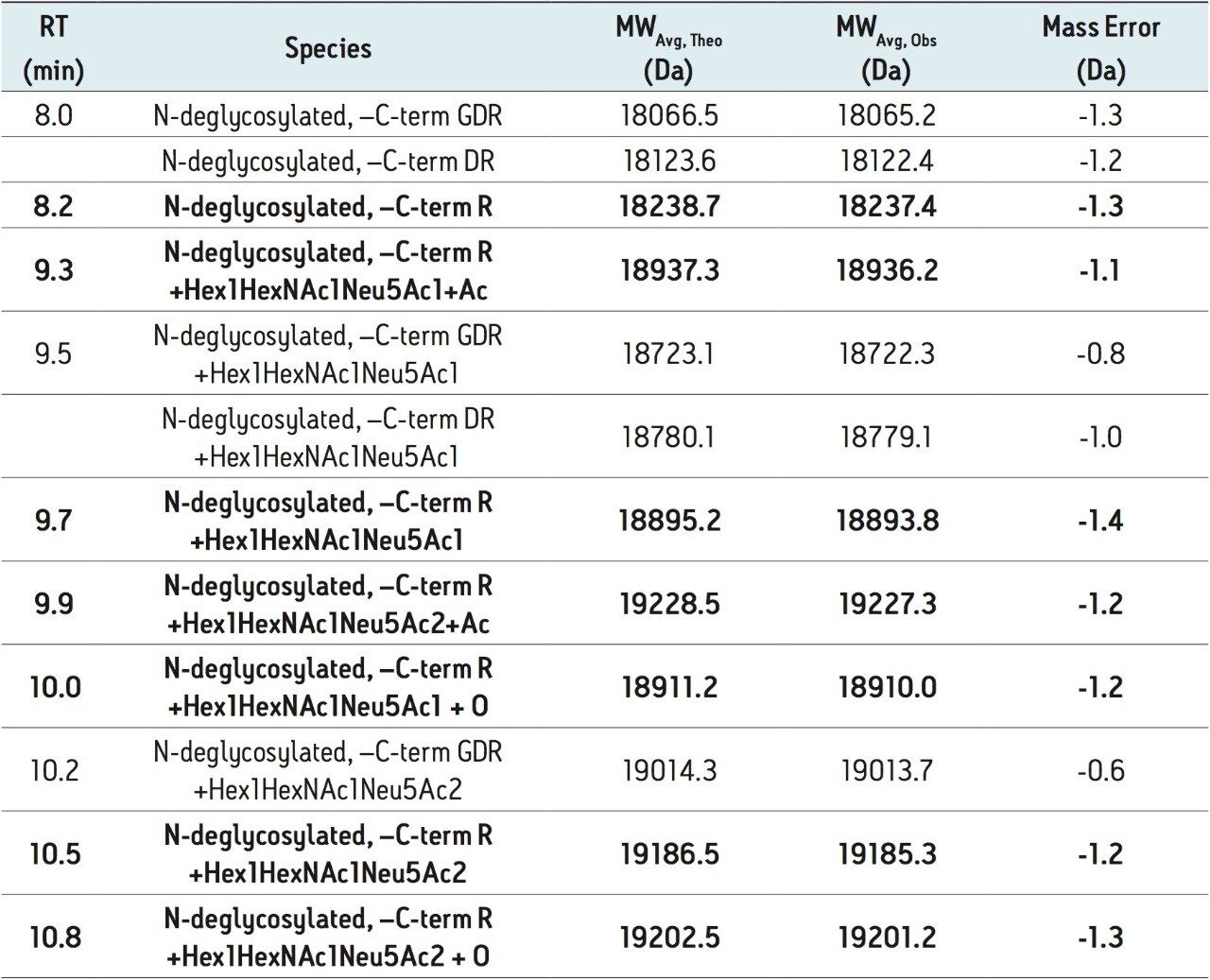
To this end, a sample of RapiFluor-MS N-glycans derived from rhEPO was profiled using HILIC. A recently introduced widepore amide column, the ACQUITY UPLC Glycoprotein BEH Amide, 300Å, 1.7 µm Column, was selected for this work to obtain high resolution N-glycan separations. This column was purposefully designed to facilitate HILIC separations of large molecules, such as glycopeptide and glycoproteins. However, the widepore particle architecture has also been shown to increase the peak capacity of highly branched, tri- and tetra-antennary N-glycans by 10–20%,17 making it an ideal choice for the HILIC profiling of EPO N-glycans, which typically exhibit high antennarity. Figure 2A shows the HILIC fluorescence and base peak intensity (BPI) MS chromatograms of the RapiFluor-MS N-glycans resulting from 0.4 µg of rhEPO. Even with this relatively limited amount of sample, high signal-to-noise chromatograms are obtained. The sensitivity of the fluorescence trace allows for accurate, relative quantitation across the profile. The signal-to-noise of the MS chromatogram is also particularly noteworthy, though it should be noted that MS sensitivity decreases as N-glycan structures become larger. Nevertheless, the quality of these particular data is made possible by use of the RapiFluor-MS reagent in combination with the Xevo G2-XS QTof, a new generation MS instrument with improved transmission efficiency and sensitivity. This QTof technology provides unprecedented sensitivity as well as high mass resolution, as can be observed in the collection of mass spectra in Figure 2B that have been used to support the assignment of various N-glycan species.