使用RapiZyme™核糖核酸酶进行可调节的RNA酶解,以确认序列并绘制修饰图谱
摘要
sgRNA和mRNA等RNA治疗药物是基因治疗应用中治疗人类疾病的重要形式。这些RNA分子的快速开发和应用需要加速开发新的分析方法。本方法旨在通过液质联用技术(LC-MS)进行寡核苷酸序列图谱分析来鉴定治疗性RNA、测定其纯度和修饰。MS测序直接检测序列中的RNA片段并定位核苷修饰。目前用于产生RNA片段的酶工具存在覆盖不完全或部分覆盖的问题,序列解析的不确定性非常高。提供具有互补剪切特异性的酶工具可以解决这个问题并填补序列空白,同时简化数据解析和提高可信度。本应用纪要评估了RapiZyme MC1和RapiZyme Cusativin的优化方案,并探讨了每种核糖核酸酶的重现性和互补特异性,重点聚焦于提高sgRNA寡核苷酸图谱分析的序列覆盖率。新工作流程采用的数据处理软件包括waters_connect™中的MAP Sequence应用程序和两款支持型MicroApp。这些软件使工作流程更加精简,可促进常规RNA片段鉴定、序列覆盖率评估和定量分析中的数据分析。
优势
- 对sgRNA实现全序列覆盖,在产生更多独特质量数的同时保持LC-MS兼容性和高信号
- 简单、可重现的方案,适用于自动化操作,无需使用化学变性剂或酶抑制剂
- 能够执行部分酶解并生成重叠酶解产物
- 在waters_connect应用程序的计算机模拟mRNA酶解计算支持下,实现基于精确质量数匹配的自动化mRNA酶解物注释
- 完全集成到waters_connect MAP Sequence中,简化数据分析
简介
背景介绍:
mRNA疫苗在应对SARS-Cov-2大流行疫情中取得的成功加速了RNA治疗药物的研发进程,这些药物不仅可用于传染病治疗,还可通过CRISPR sgRNA介导的基因编辑提供基因疗法。与此同时,分析技术的并行进步能够加速发现、开发和QC实验室中表征研究及放行检测工作的周转效率。通过LC-MS进行寡核苷酸图谱分析和测序来鉴定治疗性mRNA、确认纯度和修饰,已成为广泛使用的直接分子分析策略。与自下而上的肽图分析方法类似,RNA通过酶解产生RNA片段,然后通过液相色谱(LC)法在反相(或亲水作用)色谱柱上分离。酶解组分从色谱柱中洗脱后,经电离并进行质谱(MS)检测1。迄今为止,RNase T1是LC-MS寡核苷酸图谱分析中最常用的酶。这种酶在鸟苷残基(G)的3'-端剪切RNA。因此,RNase T1通常会产生较小的酶解产物(单体、二聚体、三聚体等),尤其是当它作用于富含G的区域时。这些产物与序列中多个位置的匹配不明确,导致数据解析模糊不清。此外,相同的RNA分子小片段不适用于计算序列覆盖率,会导致覆盖率空白和序列信息可信度低。
为了解决这些限制,沃特世针对RNA酶解开发出一组新的重组核酸内切酶,这些酶表现出与RNase T1互补的剪切特异性。重组RapiZyme MC1和RapiZyme Cusativin在识别和剪切二核苷酸位点方面表现出理想活性,如图1所示2。 此外,两种RapiZyme核糖核酸酶都有部分酶解的倾向,从而产生独特的重叠产物,适合用于LC-MS分析和序列测定。本应用纪要展示了使用RapiZyme MC1和RapiZyme Cusativin通过简单的一锅法方案对CRISPR单向导(sg)RNA实现了可重现酶解。将这些酶与IonHance™ HFIP(六氟异丙醇)和采用了mRNA Cleaver、MAP Sequence、Coverage Viewer和UNIFI处理工作流程等MicroApp的RNA数据分析工作流程结合使用,有助于加快RNA治疗药物的LC-MS表征过程3。凭借其重现性与合规架构,这些工具适用于发现实验室和QC实验室,既能支持监管申报工作,也为未来的产品放行检测提供了易于部署的解决方案。
![展示了A) RapiZyme Cusativin (Cp[A/U/G])和B) RapiZyme MC1 ([A/U/C]pU)主要剪切位置的示例图](/content/dam/waters/en/app-notes/2024/720008539/720008539en-f1.jpg.82.resize/img.jpg) 图1. 展示了A) RapiZyme Cusativin (Cp[A/U/G])和B) RapiZyme MC1 ([A/U/C]pU)主要剪切位置的示例图。“次级酶切位点”列表请参阅“结果与讨论”下的“RapiZyme核糖核酸酶的特异性”部分。
图1. 展示了A) RapiZyme Cusativin (Cp[A/U/G])和B) RapiZyme MC1 ([A/U/C]pU)主要剪切位置的示例图。“次级酶切位点”列表请参阅“结果与讨论”下的“RapiZyme核糖核酸酶的特异性”部分。
实验
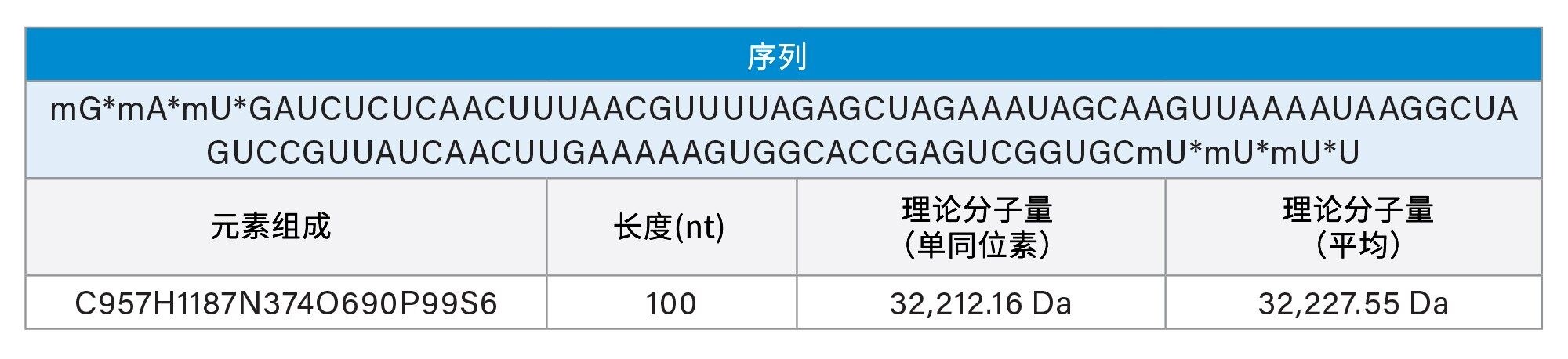 星号(*)表示PS键,“m”表示2'-O-甲基(OMe)修饰。
星号(*)表示PS键,“m”表示2'-O-甲基(OMe)修饰。
构建不含核糖核酸酶的环境:开始RNA分析操作之前,使用70%异丙醇(IPA)清洁工作区,使其不含核糖核酸酶。使用的所有试剂均不含核糖核酸酶(经认证),并用不含核糖核酸酶的水配制。
酶制备:取1瓶(10,000单位)RapiZyme MC1,溶解于200 µL 200 mM乙酸铵缓冲液中(pH 8),配制成50 U/µL的核糖核酸酶储备液;取1瓶(10,000单位)RapiZyme Cusativin,溶解于200 µL 200 mM乙酸铵缓冲液中(pH 9),配制成50 U/µL的核糖核酸酶储备液。
使用RapiZyme MC1酶解sgRNA
将100 pmol (3.2 µg) Hs.Cas9.HPRT1.2.AA sgRNA (IDT Technologies)用200 mM乙酸铵(pH 8)稀释至10 μL,在90 °C下变性2 min,然后快速冷却至4 °C。用18.2 MΩ水将1 μL RapiZyme核糖核酸酶储备液(共50单位)稀释至10 μL,然后加入各样品中,轻轻涡旋并离心以确保混合。将酶解混合物置于30 °C下温育1 h。在70 °C下温育15 min对酶进行热灭活。
使用RapiZyme Cusativin酶解sgRNA
将100 pmol (3.2 µg) Hs.Cas9.HPRT1.2.AA sgRNA (IDT Technologies)用200 mM乙酸铵(pH 9)稀释至10 μL,在90 °C下变性2 min,然后冷却至4 °C。用18.2 MΩ水将1 μL RapiZyme Cusativin储备液(共50单位)稀释至10 μL,然后加入各样品中,轻轻涡旋并离心以确保混合。将酶解混合物置于30 °C下温育1 h。在75 °C下温育15 min对酶进行热灭活。
使用RNase T1酶解sgRNA
将100 pmol (3.2 µg) Hs.Cas9.HPRT1.2.AA sgRNA (IDT Technologies)用200 mM乙酸铵(pH 7)稀释至10 μL,在90 °C下变性2 min,然后冷却至4 °C。加入用10 μL 18.2 MΩ水稀释的250单位RNase T1 (Worthington Biochemical Corporation),轻轻涡旋并离心以确保混合。将酶解混合物置于37 °C下温育1 h。在80 °C下温育15 min对酶进行热灭活。
使用RNase 4酶解sgRNA
将6.25 μL 10 µg Hs.Cas9.HPRT1.2.AA sgRNA (IDT Technologies)与3.75 μL 8 M尿素混合,得到最终浓度为3 M尿素的溶液(NEB推荐的浓度)。在90 °C下使RNA变性5 min,然后冷却至25 °C。按照NEB的建议,用20 μL 1.5 X NEBuffer™ r1.1将变性的RNA混合物稀释至最终体积30 µL,并加入1 μL RNase 4 (NEB #M1284)储备液(50 U/μL)。将混合物置于37 °C下温育1小时。加入1 μL NEB核糖核酸酶抑制剂(NEB #M0314)终止反应,然后在室温下温育10 min。
随后将各酶解混合物转移至低结合性QuanRecovery™样品瓶(P/N:186009186)中,对各种酶进样等量的RNA酶解物(50 pmol或1.6 µg当量)进行LC-MS分析。
液相色谱条件
|
色谱柱: |
ACQUITY™ Premier BEH™ C18寡核苷酸分析专用柱, 300 Å, 1.7 µm, 2.1 x 50 mm(P/N:186010539) |
|
流动相A: |
0.1% N,N-二异丙基乙胺(DIPEA)和1% (v/v) IonHance HFIP(P/N:186010781),溶于18.2 MΩ水 |
|
流动相B: |
0.0375% DIPEA和0.075% (v/v) IonHance HFIP(P/N:186010781),溶于55:10:35乙腈:甲醇:18.2 MΩ水 |
|
液相色谱系统: |
ACQUITY Premier BSM系统 |
|
检测器: |
Xevo G3 QTof |
|
波长: |
254 nm |
|
流速: |
0.4 mL/min |
|
进样: |
10 µL |
|
柱温: |
70 °C |
|
样品温度: |
4 °C |
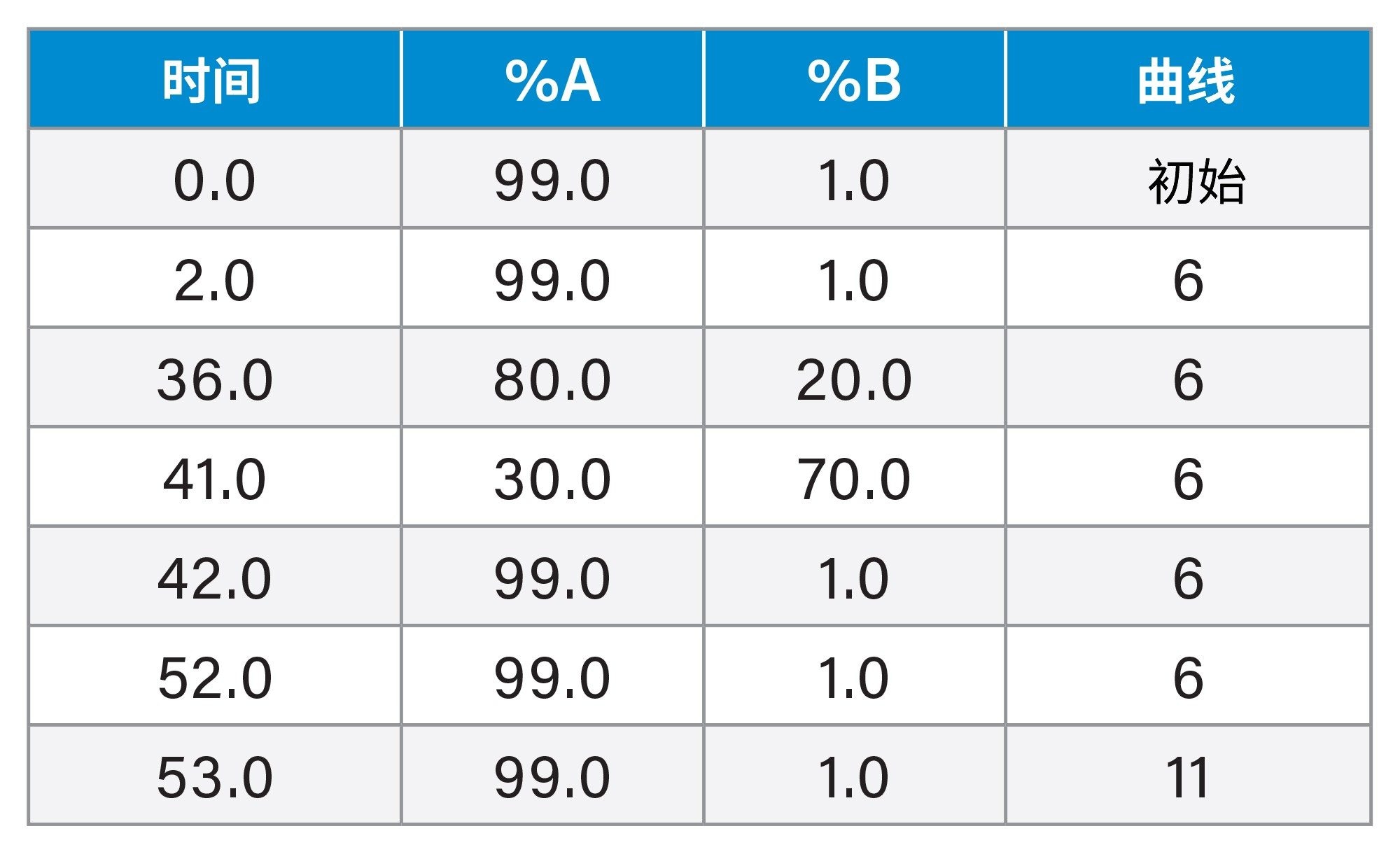
梯度表
质谱条件
|
离子源类型: |
ESI |
|
极性: |
负 |
|
分析器模式: |
灵敏度 |
|
毛细管电压: |
1.5 kV |
|
样品锥孔电压: |
40 V |
|
离子源温度: |
100 °C |
|
脱溶剂气温度: |
550 °C |
|
锥孔气流速: |
50 L/h |
|
脱溶剂气流速: |
650 L/h |
|
扫描质量数范围: |
550~2000 |
|
扫描时间: |
1.0 s |
|
碰撞能量: |
低,6 V |
|
高能阶梯: |
10~45 V |
|
智能数据捕获 - 强度阈值: |
中(10) |
|
锁定校正: |
自动(间隔30秒) |
|
事件:LC,样品: |
2.0~40.0 min |
|
锁定质量数:亮氨酸脑啡肽 |
554.26202 (M-H+)1- |
|
锁定质量数:合并宽度和质量数窗口: |
3次扫描,0.5 m/z |
结果与讨论
本研究的目的是评估RapiZyme核糖核酸酶在RNA酶解中的批次间性能,及其在治疗性RNA寡核苷酸图谱分析中的实用性。在本研究中,我们考察了用于靶向人HPRT(次黄嘌呤鸟嘌呤磷酸核糖转移酶)基因(Hs.Cas9.HPRT1.2.AA)的CRISPR sgRNA的分析结果。
RapiZyme核糖核酸酶的特异性
RapiZyme MC1和Cusativin在特定的二核苷酸位点表现出对底物对接和切割的偏好,如表1所示。RapiZyme MC1对RNA的剪切位点为尿苷5'-端的ApU、CpU和UpU处,同时对CpA和CpG二核苷酸组合的偏好性略低。此外,MC1不会剪切GpU键。RapiZyme Cusativin表现出稍微更宽的特异性,对RNA的剪切位点为胞苷3'-端的CpA、CpG、CpU处,同时对ApU、GpU、UpU和UpA键的偏好性略低。此外,Cusativin几乎或完全不会剪切CpC键。尽管这些酶的特异性稍宽,但能够可重现地实现部分酶解,并且不需要使用保护性杂化寡核苷酸就能实现这样的结果。这一能力使其有可能生成具有重叠特征的酶解产物,从而提高序列覆盖率和促进可靠的数据解析。通过漏切实现部分酶解是RapiZyme MC1和Cusativin的特有功能,因此能够产生更多样的酶解组分,从而为研究RNA分析物的序列组分和结构细节提供更多机会。这些较长的酶解产物生成的质量数值降低了检出异构体的频率,因此不再需要更严格的MS/MS分析和序列解析。
用于数据解析的信息学软件:Waters MicroApp mRNA Cleaver可根据常见酶(包括RapiZyme核糖核酸酶)的酶切特异性预测其预期酶解产物的模拟质量数值。此文件可用于通过waters_connect应用程序的MAP Sequence来匹配和鉴定LC-MS分析中观察到的酶解产物。使用这些结果计算序列覆盖率,并使用Coverage Viewer将唯一和非唯一酶解产物的序列覆盖率可视化。这些功能在当前使用HPRT sgRNA作为底物的RNA分析物研究中进行了介绍。
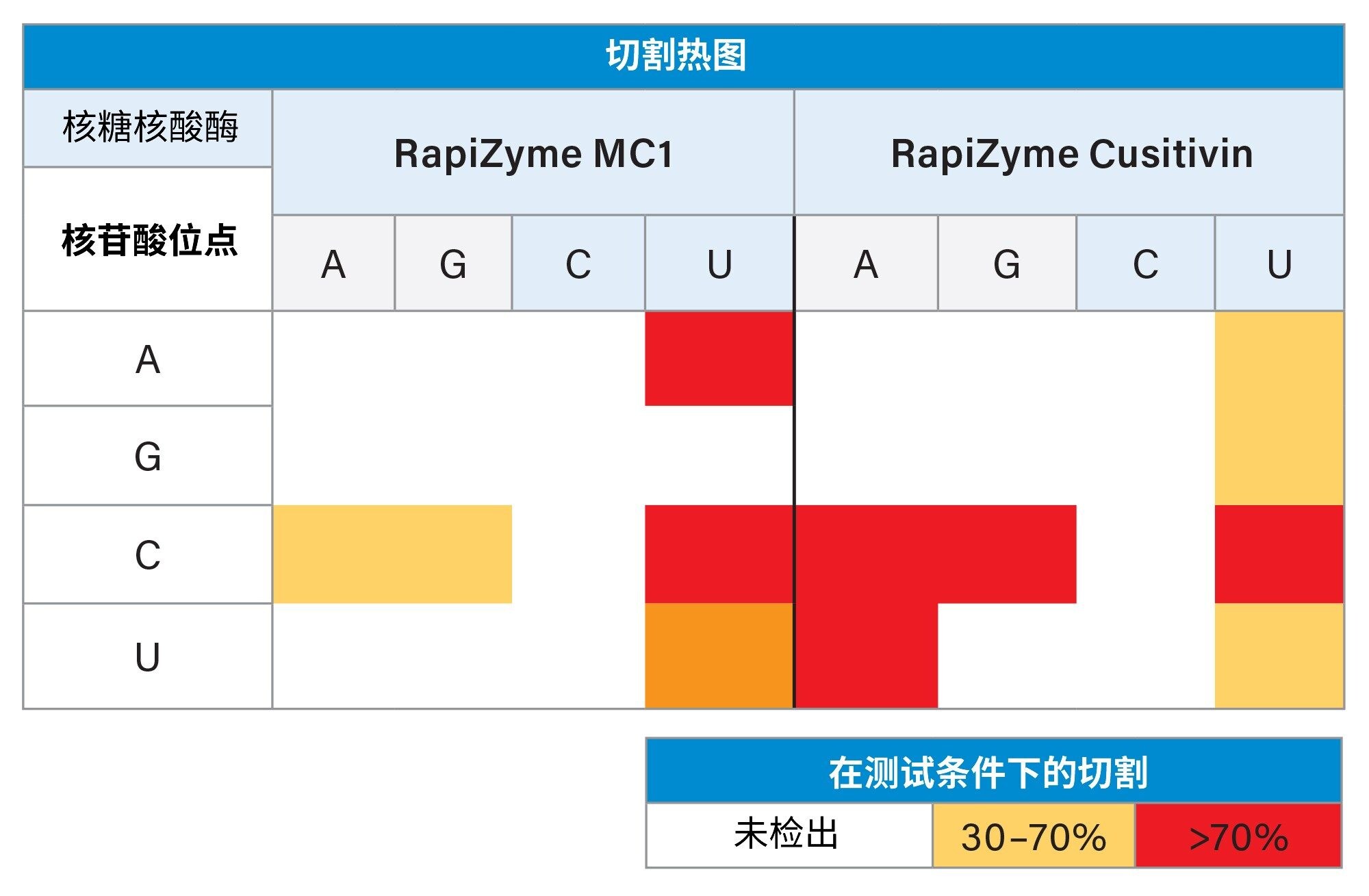 表1.RapiZyme MC1和RapiZyme Cusativin对每种二核苷酸组合的相对剪切反应速率近似值
表1.RapiZyme MC1和RapiZyme Cusativin对每种二核苷酸组合的相对剪切反应速率近似值
RapiZyme核糖核酸酶的批次间重现性
在寡核苷酸图谱分析的样品前处理中,RNA底物的可重现酶解至关重要。我们在多个批次中使用HPRT sgRNA对RapiZyme MC1和RapiZyme Cusativin的可重现酶解行为进行了三重评估。图2展示了酶解物的LC-UV-MS分析结果。两种酶的UV谱图均表现出优异的重现性,峰高差异极小,但RapiZyme MC1的酶解产物子集在丰度方面存在细微差异(图2A和2B)。此外,本研究使用MAP Sequence应用程序评估了MS信号,以根据mRNA Cleaver的输入值鉴定酶解产物及其保留时间。使用mRNA Cleaver生成含4个漏切位点的模拟预测酶解产物。利用创建的酶解产物文件和MAP Sequence应用程序,根据LC-MS数据鉴定酶解产物。对MAP Sequence输出中的所有备选分配进行手动确证后,制备UNIFI Excel模板文件,注明寡核苷酸酶解产物、元素组成和保留时间。为展示和可视化重现性,我们将该文件作为谱库组件导入waters_connect管理工具,以便对所有进样使用UNIFI数据分析处理方法。根据MS分析的结果,在查看模式下绘制提取离子流色谱图叠加图。RapiZyme Cusativin和RapiZyme MC1的叠加图分别如图2C和2D所示。
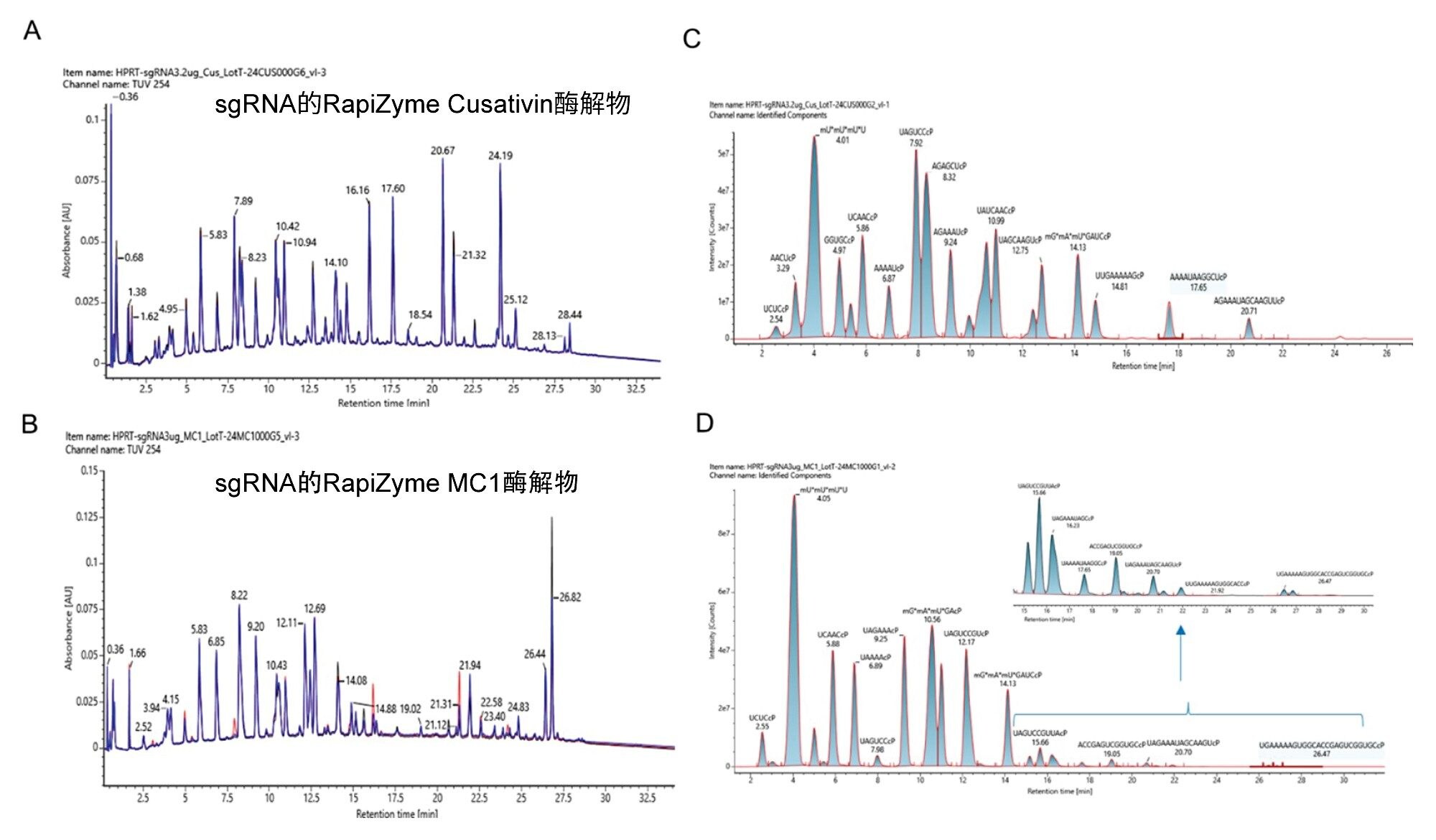 图2.用重组RapiZyme Cusativin和MC1酶解HPRT sgRNA进行的LC-UV-MS分析。使用三个独立批次的重组酶RapiZyme Cusativin (A)和RapiZyme MC1 (B)酶解HPRT sgRNA,将三张UV色谱图(黑色、蓝色和红色迹线)相互叠加,以展示三种制剂的可重现酶解行为。(C) RapiZyme Cusativin酶解产物鉴定的叠加提取离子流色谱图,在代表性TUV迹线中显示了丰度。(D) RapiZyme MC1酶解产物鉴定的叠加提取离子流色谱图,在代表性TUV迹线中显示了丰度。插图显示了RapiZyme MC1酶解产物的低丰度酶解产物谱图。
图2.用重组RapiZyme Cusativin和MC1酶解HPRT sgRNA进行的LC-UV-MS分析。使用三个独立批次的重组酶RapiZyme Cusativin (A)和RapiZyme MC1 (B)酶解HPRT sgRNA,将三张UV色谱图(黑色、蓝色和红色迹线)相互叠加,以展示三种制剂的可重现酶解行为。(C) RapiZyme Cusativin酶解产物鉴定的叠加提取离子流色谱图,在代表性TUV迹线中显示了丰度。(D) RapiZyme MC1酶解产物鉴定的叠加提取离子流色谱图,在代表性TUV迹线中显示了丰度。插图显示了RapiZyme MC1酶解产物的低丰度酶解产物谱图。
为了比较RapiZyme Cusativin和MC1与RNase T1和RNase 4的酶解产物谱图,按照方法部分所述的方法,用这四种核糖核酸酶酶解sgRNA。四种核糖核酸酶的总离子流色谱图(TIC)如图3所示。与RNase T1和RNase 4相比,在RapiZyme MC1和Cusativin的酶解物中可以轻松观察到种类丰富、MS信号丰度高的酶解产物。此外,即使以相似的载样量(50 pmol当量酶解物)进样分析,RNase 4的酶解物信号强度也较弱。
RNase T1和RNase 4酶解物的LC-MS数据分析表明,存在一组离散的酶解产物,即使mRNA Cleaver Microapp在这些酶的酶解产物预测中考虑了漏切,这些产物也没有重叠特征。将在每种酶中观察到的酶解产物分别叠加到HPRT sgRNA序列上(如图4所示)进行比较。
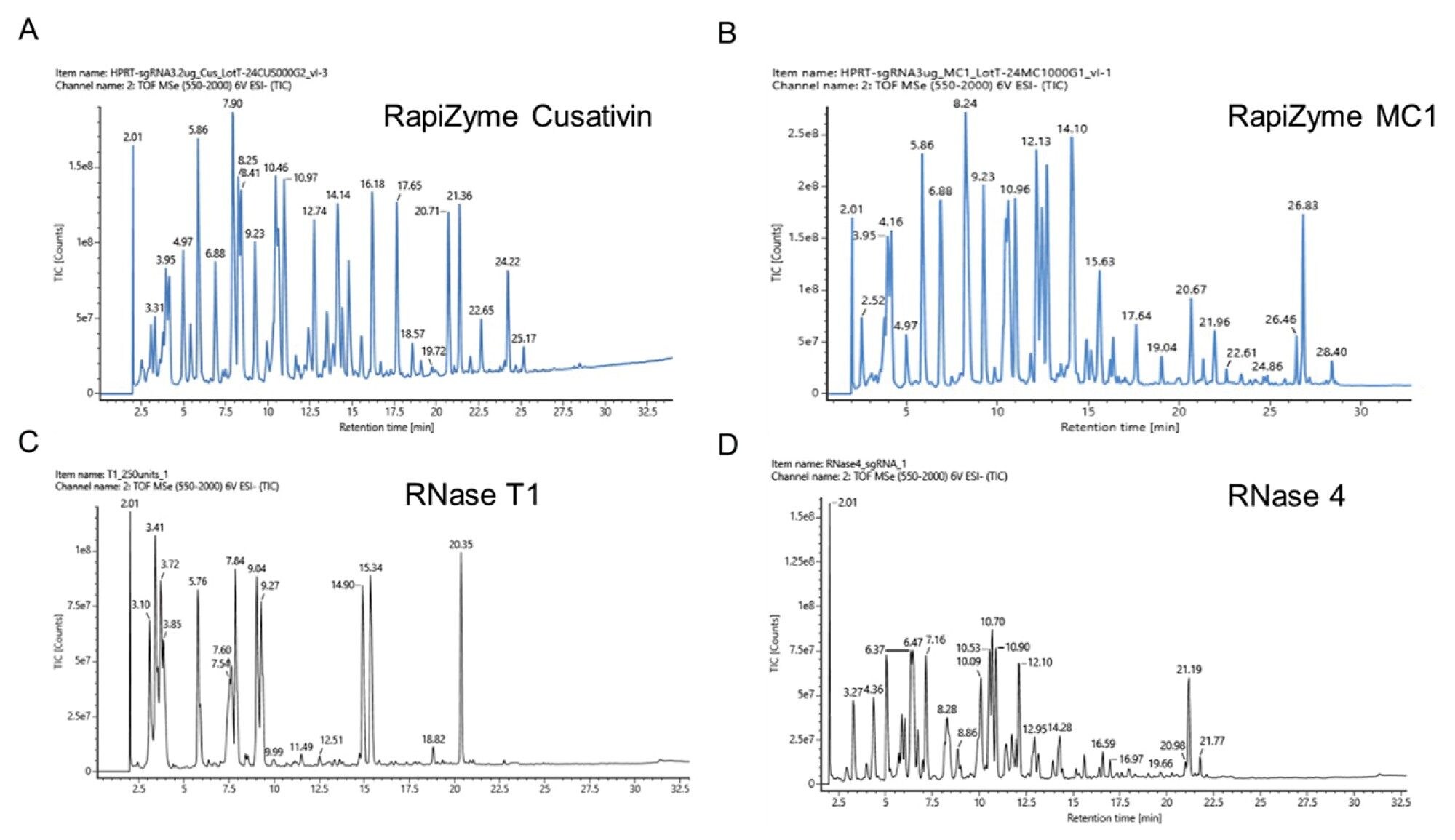 图3.RapiZyme Cusativin (A)、RapiZyme MC1 (B)、RNase T1 (C)和RNase 4 (D)的HPRT1 sgRNA酶解物LC-MS分析结果。
图3.RapiZyme Cusativin (A)、RapiZyme MC1 (B)、RNase T1 (C)和RNase 4 (D)的HPRT1 sgRNA酶解物LC-MS分析结果。
RapiZyme MC1和Cusativin酶解产物均出现重叠,提供了额外的序列位置和背景,从而提高了数据解析的可信度。这些序列重叠可能是由于RapiZyme MC1和Cusativin能够对RNA实现可重现的部分酶解。此外,将在RapiZyme MC1和Cusativin中观察到的酶解产物映射到sgRNA序列时,实现了全(100%)序列覆盖。相比之下,RNase T1只能实现部分序列覆盖(约78%),存在覆盖率差距和不确定性,可能将同一种酶解产物映射到至少两个序列区域。尽管RNase 4酶解物实现了全序列覆盖,但其酶解产物较为分散,没有序列重叠。
关于酶解产物上的末端磷酸酯类型,RapiZyme MC1和Cusativin酶解物主要为环状磷酸酯,而RNase 4为等量的环状和线性磷酸酯,这有助于解释在RNase 4数据中观察到的低信号强度。而线性磷酸酯是RNase T1酶解产物的主要形式(表2)。酶解产物的长度也各不相同,与其他酶相比,RapiZyme MC1的酶解产物最长(27 nt)。
 图4.RNase T1、RNase 4、RapiZyme MC1和RapiZyme Cusativin的HPRT sgRNA酶解产物鉴定示意图。粗体碱基表示在重叠酶解产物中检测到残基的序列区域。以p结尾的序列(例如AACGUUUUp)表示检测到高丰度的线性磷酸酯产物。
图4.RNase T1、RNase 4、RapiZyme MC1和RapiZyme Cusativin的HPRT sgRNA酶解产物鉴定示意图。粗体碱基表示在重叠酶解产物中检测到残基的序列区域。以p结尾的序列(例如AACGUUUUp)表示检测到高丰度的线性磷酸酯产物。
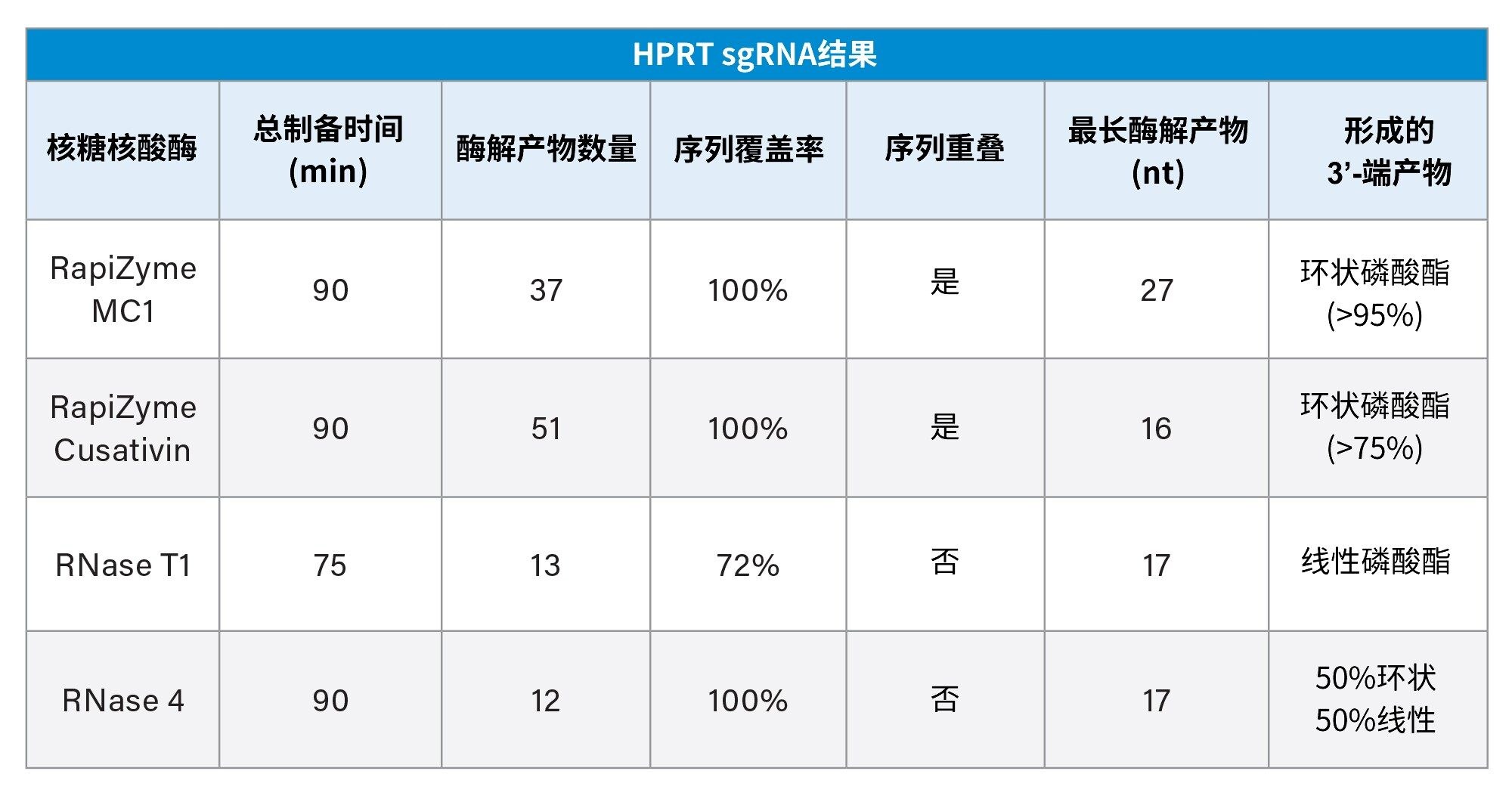 表2.HPRT sgRNA结果
表2.HPRT sgRNA结果
结论
本研究建立了一个稳定的分析工作流程,包括使用IP-RP-LC-MS研究合成RNA治疗药物的样品前处理和数据分析工作。
- 使用RapiZyme MC1和RapiZyme Cusativin对合成sgRNA实现了可重现地酶解,样品前处理方案中消耗的RNA仅100 pmol或3.2 µg,并且分析时仅进样50 pmol (1.6 µg) RNA酶解物
- 使用ACQUITY Premier二元系统、ACQUITY Premier BEH C18寡核苷酸分析专用柱和IonHance HFIP流动相试剂进行离子反相色谱分析,短链和长链酶解产物均获得了较高的色谱分离度以及优异的UV和MS信号
- 本研究基于精确质量数匹配鉴定RNA酶解物片段并绘制图谱,使用mRNA Cleaver、MAP Sequence和Coverage Viewer的计算机模拟RNA酶解计算发挥了助力
- 两种RapiZyme核糖核酸酶均获得了完全覆盖(存在序列重叠),从而提供了可靠的数据解析和输出
mRNA疫苗、CRISPR sgRNA以及合成寡核苷酸等创新、更先进的RNA治疗药物正处于迅猛发展阶段。为解决与表征相关的复杂问题,需要使用新的样品前处理、色谱分离以及数据分析和数据解析信息学解决方案。本应用纪要中概述的方法证明,使用RapiZyme MC1和RapiZyme Cusativin,通过生成独特的酶解产物和LC-MS分析,能够可重现地表征和监测RNA治疗药物(例如sgRNA),从而实现可靠、完全的序列覆盖。本研究证明,通过采用具有不同剪切偏好的多种核酸酶可以生成正交和附加的序列图谱信息。
有关用于RNA表征的waters_connect MAP Sequence工作流程的详细信息,请参阅《使用集成式UPLC-MS和信息学工作流程进行RNA酶解物图谱分析》(RNA Digestion Product Mapping Using an Integrated UPLC-MS and Informatics Workflow),沃特世应用纪要 - 720008553。
参考资料
- Gau, B.; Dawdy, A.; Wang, H.L. et al. (2023) Oligonucleotide Mapping Via Mass Spectrometry to Enable Comprehensive Primary Structure Characterization of an mRNA Vaccine Against SARS-Cov-2.Scientific Reports 13, 9038.
- Thakur P, Atway J, Limbach PA, Addepalli B. RNA Cleavage Properties of Nucleobase-Specific RNase MC1 and Cusativin are Determined by the Dinucleotide-Binding Interactions in the Enzyme-Active Site. Int J Mol Sci. 2022 Jun 24;23(13):7021.
- Gaye MM, Fox J, Vissers JPC, Reah I, Knowles C, Lauber M (2022) 使用离子对液相色谱和质谱法进行合成mRNA寡核苷酸图谱分析.沃特世应用纪要 – 720007669ZH.2022年6月.
720008539ZH,2024年9月